THE ROLE OF ENDOTHELIAL PROGENITOR CELLS ON VASCULAR REGENERATION PROCESS
on
PERAN SEL PROGENITOR ENDOTEL DALAM PROSES REGENERASI VASKULAR
*Katrin, **Desak Made Wihandani
*Mahasiswa Semester XI
Fakultas Kedokteran Universitas Udayana, Denpasar, Bali, 80232
**Dosen Pembimbing Bagian Biokimia
Fakultas Kedokteran Universitas Udayana, Denpasar, Bali, 80232
ABSTRAK
Latar belakang: Sel progenitor endotel (EPC) memiliki potensi yang sangat besar dalam meregenerasi pembuluh darah melalui mekanisme angiogenesis, vaskulogenesis, dan neovaskularisasi, terutama reperfusi setelah terjadi iskemia. Namun demikian, hingga saat ini belum terdapat konsensus universal mengenai definisi EPC karena penelitian berbasis biologi seluler yang beragam.
Tujuan: Tujuan dari tinjauan pustaka ini adalah untuk menguraikan klasifikasi subtipe EPC. Selain itu, tinjauan pustaka ini juga menjelaskan tentang Sejumlah sitokin dan faktor pertumbuhan diketahui dapat menginduksi ekspresi reseptor serta, migrasi, adhesi, invasi, dan diferensiasi EPC menjadi sel endotel dewasa.
Metodologi: Tinjauan pustaka ini merangkum subtipe klasifikasi EPC berdasarkan penelitian terbaru, selain peranannya dalam neovaskularisasi dan regenerasi sel endotel melalui literatur penemuan dan sintesis, serta membandingkan berbagai artikel penelitian.
Hasil: Terdapat beberapa strategi untuk mengklasifikasikan EPC berdasarkan beberapa kriteria meliputi evaluasi adhesi secara in vitro, antigen penanda permukaan, morfologi seluler, karakteristik khusus (non-fagosit), dan kemampuan membentuk formasi tabung in vitro dan pembuluh darah in vivo. EPC terutama berasal dari sumsum tulang dan dimobilisasi saat iskemia dan inflamasi. Beberapa sitokin dan faktor pertumbuhan diketahui menyebabkan terjadinya mobilisasi tersebut.
Kesimpulan: EPC mempunyai peranan yang sangat penting dalam proses neovaskularisasi dan angiogenesis pada saat terjadi iskemia atau inflamasi. Definisi yang mencerminkan identitas EPC belum ada yang tepat hinggal saat ini. Identitas tersebut harus tepat dan dirangkum dalam suatu konsensus internasional sebelum melangkah lebih jauh dalam strategi terapi yang lebih rumit dengan memanfaatkan teknologi sel.
Kata kunci: EPC, CD34, neovaskularisasi, regenerasi endotelium, identitas seluler
THE ROLE OF ENDOTHELIAL PROGENITOR CELLS ON VASCULAR REGENERATION PROCESS
ABSTRACT
Background: Endothelial progenitor cells (EPCs) possesses an enormous potential in regenerating blood vessels and maintain its integrity and elasticity through angiogenesis, vasculogenesis, and neovascularization, mainly during reperfusion after ischemia. However, to date, there has been no universal consensus regarding the precise definition of EPCs due to highly variable cell-based research in this growing field.
Aim of study: This review aims to elucidate the various approach to classify EPC subtypes. Moreover, we also elaborate the EPCs signaling mechanism, comprising the cytokines and growth factors involved in EPCs receptor expression, migration, adhesion, invasion, and differentiation process into mature endothelial cells.
Method: This review summarizes the contemporary EPCs subtype classification as well as their role in neovascularization and endothelial regeneration through extensive literature discoveries and synthesis, comprising mainly research articles.
Results: EPCs classification can be derived from several approaches, including in vitro adhesion assay, antigen surface markers, cellular morphology, specific characteristics (non-ingestion), and ability to form tube in vitro and blood vessels in vivo. EPCs mainly are derived bone marrow and mobilized during ischemia and inflammation. A number of cytokines and growth factors have been known to induce.
Conclusion: EPCs play a crucial role in neovascularization and angiogenesis process in times of ischemia or inflammation. However, to date, there has been no precise definition regarding EPCs identity. Its identity must be ideally confirmed and unified in a single international consensus before stepping further into a more sophisticated therapeutic strategies by utilizing these cell-based technology.
Keywords: EPCs, neovascularization, endothelial regeneration, cellular identity
PENDAHULUAN
Sel endotel memiliki peran penting dalam mempertahankan integritas vaskuler.1 Melalui formasi khusus, sel endotel melapisi seluruh pembuluh darah di dalam tubuh. Dalam kondisi normal, endotel vaskuler membentuk permukaan non-adhesif yang impermeabel terhadap leukosit dan platelet sehingga mencegah terjadinya formasi trombus.2 Selain itu, untuk menjaga elastisitas vaskuler, sel endotel mensekresikan nitrit oksida (NO) yang berfungsi sebagai vasodilator dan merelaksasikan otot polos yang terdapat pada dinding vaskuler.3 Pada kasus jejas vaskuler (misal karena iskemia atau
infark), endotel vaskuler juga berperan dalam neovaskularisasi untuk meningkatkan
reperfusi ke daerah perlukaan sehingga mempercepat penyembuhan.1,4
Hingga saat ini masih terjadi perdebatan mengenai jenis sel endotel yang terlibat dalam neovaskularisasi ketika terjadi jejas vaskuler. Awalnya, proses angiogenesis dianggap sebagai pemanjangan sel endotel dewasa dari pembuluh darah yang telah ada.1,5 Namun terdapat indikasi adanya sel unipoten beredar dalam sirkulasi darah yang bersifat klonogenik (mampu memperbarui diri), proliferatif, dan mampu berdiferensiasi menjadi sel endotel dewasa.6 Sel-sel tersebut sensitif terhadap sitokin proinflamasi, 7-faktor pertumbuhan (growth factor), dan hipoksia yang terjadi saat inflamasi vaskuler.7-10 Saat terstimulasi, sel-sel ini bermigrasi menuju area jejas, berdiferensiasi menjadi sel endotel dewasa dan menggantikan endotel yang rusak. Setelah berhasil diisolasi oleh Asahara et al pada tahun 1997, sel tersebut diberi nama sel progenitor endotel (endothelial progenitor cell/EPC).11
Investigasi EPC dalam satu dekade terakhir relatif signifikan. Melalui lebih dari 7600 penelitian12 yang telah dilakukan sejak penemuan Asahara et al11, beberapa aspek penting mengenai karakterisasi, jalur pensinyalan, faktor pro- dan kontra-mobilisasi, serta peranan EPC dalam regenerasi vaskuler mulai dapat didefinisikan secara konkret. Namun layaknya sebuah penemuan baru, berbagai riset EPC tidak selalu saling mendukung teori yang dikemukakan. Akibatnya, telaah referensi berdasarkan beberapa bukti ilmiah saja tidak dapat menggambarkan perkembangan riset EPC secara komprehensif. Oleh karena itu, artikel ini akan berusaha memaparkan sejumlah aspek penting yang berkaitan dengan peranan EPC dalam proses regenerasi vaskuler.
PEMBAHASAN
Definisi dan Karakterisasi EPC
EPC didefinisikan sebagai sel unipoten yang bersifat klonogenesis (dapat memperbarui diri/self renewal), proliferatif, dan dapat berdiferensiasi menjadi sel endotel dewasa menggantikan endotel vaskuler yang nekrosis atau apoptosis.2,5,11,12 Namun demikian, EPC dilaporkan bersifat multipoten karena dapat ber-transdiferensiasi (diferensiasi menjadi sel yang berbeda karakteristik fenotipe, morfologi, dan fungsional) menjadi kardiomiosit.13 Identifikasi EPC menjadi sulit dan beragam karena hingga saat ini belum terdapat penanda permukaan (surface marker) khas yang dapat membedakan EPC dari turunan sel lain, seperti sel punca hematopoietik (hematopoietic stem cell/HSC) dan prekursor myeloid. Oleh karena itu, para ahli berusaha mengklasifikasikan berbagai temuan yang diduga EPC melalui penanda permukaan yang paling sering diekspresikan, sifat adhesi in vitro, morfologi, kemampuan absorbsi penanda spesifik (mis. acetylated low density lipoprotein/acLDL), formasi tabung in vitro, dan formasi pembuluh darah in vivo.5,12,14
Asahara et al11 pertama kali mengisolasi sel yang mengekspresikan antigen cluster of differentiation 34+ (CD34+) dari sirkulasi darah. CD34 sebelumnya diketahui sebagai penanda molekuler sel hematopoietik.15 Ketika ditempatkan pada petri berlapis fibronektin, sel CD34+ mampu melekat (adhesi) pada cawan dan membentuk kumpulan sel bundar pada bagian sentral dan sel yang menonjolkan formasi spindel pada daerah perifer (splindle-shaped cells). Sel tersebut bila diinjeksikan pada hewan coba dengan iskemia hindlimb mengekspresikan VEGFR2 (Vascular Endothelial Growth Factor Receptor 2) (disebut juga FDR pada manusia dan fetal liver kinase-1/Flk-1 pada mencit)
dan terlokalisir pada daerah neovaskularisasi. Sel ini kemudian diberi nama endothelial cell colony-forming units (CFU-EC).11
Penelitian Asahara et al11 dilanjutkan oleh Ito et al16 dengan sedikit modifikasi. Sel CD34+ diisolasi dari sel-sel mononuklear darah tepi dan ditempatkan pada petri berlapis fibronektin. Setelah 24 jam, sel-sel yang tidak melekat (adhesi) pada petri dipindahkan pada petri baru yang juga berlapis fibronektin. 7 hari kemudian, terlihat kumpulan sel dengan peningkatan ekspresi CD31, CD34, VEGFR2, Tyrosine kinase 2 (Tie-2) dan E selectin, serta ekspresi mRNA eNOS yang diberi nama EPC-derived colonies. Prosedur awal penelitian ini dengan membuang sel-sel yang mudah adhesi bertujuan mengeksklusi monosit, makrofag, dan sel endotel dewasa yang beredar dalam sirkulasi darah, hingga menyisakan EPC yang diduga memiliki kemampuan adhesi lebih lambat dibandingkan ketiga sel tersebut.12,16
Modifikasi terakhir dilakukan oleh Hill et al17, yakni dengan menempatkan sel CD34+ pada petri fibronektin selama 48 jam dan mengkultur ulang sel-sel non-adhesi pada fase pertama menggunakan medium yang sama. Formasi koloni yang dihasilkan memiliki penanda permukaan VEGFR2, CD31, Bandeiraea Simplicifolia (BS-1) lectin dan menyerap ac-LDL, kemudian diberi nama colony-forming unit-Hill (CFU-Hill). Hill et al17 menunjukkan bahwa jumlah CFU-Hill berkurang pada pasien diabetes, hipertensi, dan peningkatan serum kolesterol. Selain itu, juga dijumpai korelasi berkebalikan antara jumlah CFU-Hill yang bersirkulasi dalam darah tepi dan skor risiko Framingham yang kemudian digunakan sebagai prediktor risiko kardiovaskuler.5
Ketiga studi menyimpulkan bahwa angioblas (termasuk EPC) memiliki antigen permukaan yang sama dengan HSC (CD34) namun mampu berdiferensiasi menjadi sel endotel dan beperan penting dalam neovaskularisasi. Namun studi lebih lanjut tidak
dapat membuktikan kemampuan CFU-EC, EPC-derived colonies, dan CFU-Hill dalam
membentuk formasi tabung in vitro, maupun formasi pembuluh darah in vivo.
Vasa et al19,20 menawarkan isolasi jenis sel yang berbeda, walaupun dikultur berbasis prosedur yang sama dengan ketiga penelitian sebelumnya. Sel-sel monuklear dari darah tepi ditempatkan pada petri berlapis fibronektin dan medium pertumbuhan sel endotel. Setelah 4 hari, sel-sel yang tidak melekat dibuang sedangkan sel-sel yang adhesi diuji kemampuannya untuk mengabsorbsi ac-LDL dan berikatan dengan label fluoresen Ulex europaeus aglutinin 1 (UEA-1). Ternyata, sel tersebut dapat menyerap kedua antigen secara optimal, seperti halnya sel endotel dewasa yang juga memiliki ciri tersebut. Kumpulan sel ini mengekspresikan antigen permukaan yang serupa dengan EPC yang diidentifikasi pada tiga penelitian sebelumnya, yakni VEGFR2, von Willdebrand Factor (vWF), E-selectin, dan mRNA eNOS. Penelitian lanjutan juga mengonfirmasi kemampuan sel-sel ini dalam proses angiogenesis, sehingga disimpulkan bahwa monosit dapat ber-transdiferensiasi menjadi EPC.
Namun seperti yang dikhawatirkan Ito et al16, sel yang menunjukkan sifat adhesi segera pada medium kultur dicurigai sebagai monosit atau makrofag. Dugaan ini ternyata terbukti dengan ditemukannya ekspresi antigen CD45, CD11b-c,dan CD14 yang juga dimiliki oleh sel monosit dan makrofag. Lebih lanjut, sel-sel tersebut mengabsorpsi antigen asing secara aktif, seperti ciri khas monosit dan makrofag yang tidak dijumpai pada sel endotel dewasa. Metode isolasi sel yang mengadakan adhesi spontan dengan fibronektin telah dijelaskan sebagai prosedur isolasi sel monosit yang efisien (>90% sel yang tersisolasi adalah monosit).21 Sedangkan, peranan sel-sel monosit dalam angiogenesis dapat dijelaskan sebagai efek parakrin berupa sekresi sitokin yang berfungsi menarik EPC untuk bermigrasi ke daerah lesi (misal monocyte chemotactic
protein/MCP-1), dan faktor pertumbuhan (colony forming unit-granulocyte
monocyte/CFU-GM, dan colony forming unit-granulocyte/CFU-G).22,23 Selain itu, sangat memungkinkan terdapat makrofag yang melapisi area vaskuler “kosong” karena sel endotel di daerah tersebut telah terdenudasi, sehingga kontak langsung makrofag dengan sirkulasi darah menimbulkan proses adaptasi sehingga sel tersebut mengekspresikan antigen dan berfungsi layaknya sel endotel dewasa yang pada kenyataannya tidak pernah ber-transdiferensiasi menjadi sel endotel murni (dengan kehilangan fenotipe dan fungsi makrofag).12 Hal tersebut telah dibuktikan dengan fenomena fagositosis antigen India ink dan bakteri E.coli pada sel kultur yang mengekspresikan antigen permukaan sel endotel.18,24
Kumpulan sel ketiga berhasil diisolasi oleh Lin et al.25 Sel ini diisolasi dari darah tepi pasien yang telah menjalani transplantasi sumsum tulang. Sel tersebut kemudian dikultur pada petri berlapis kolagen (bukan fibronektin) dan ditambahkan medium kultur endothelial growth media (EGM-2). Berbeda dengan prosedur grup pertama, sel non-adheren (yang tidak melekat pada petri) dibuang beserta medium kulturnya, dan ditambahkan EGM-2 yang baru pada sisa sel yang adhesi pada petri. Hal tersebut dilakukan secara periodik hingga kemunculan koloni sel yang mampu berproliferasi optimum pada hari ke-14 hingga 21, sehingga diberi nama endothelial colony forming cell (ECFC).18,25 ECFC mengekspresikan antigen permukaan yang identik dengan sel endotel dewasa, seperti CD31, CD34, CD105, vWF, dan VEGFR2 (KDR), namun hanya mengekspresikan antigen permukaan monosit/makrofag (CD45, CD14, CD115) dalam jumlah sangat kecil. Sedangkan CFU-EC hasil isolasi Asahara et al.11 mengekspresikan antigen makrofag secara prominen (lihat perbandingan antigen permukaan CFU-EC dan ECFC pada Tabel 1). ECFC tidak memfagosit bakteri (E.coli)
seperti halnya sel endotel dewasa. Selain itu, ECFC positif terhadap UEA-1 dan Ac-LDL, serupa dengan sel endotel.
Yoder et al.18 menganalisis secara head-to-head mengenai penanda permukaan, klonogenesis, dan fungsionalitas CFU-EC terhadap ECFC yang menghasilkan bukti dan kesimpulan definitif mengenai kemiripan ECFC terhadap sel endotel dewasa, dibandingkan CFU-EC. Dalam penelitiannya, Yoder et al. mengisolasi kedua tipe sel dari subjek yang sama, yakni menderita polisitemia vera dengan mutasi pada gen Janus Activating Kinase 2 (JAK2) V617F (subtitusi as.amino valin menjadi fenilalanin pada kodon 617) yang diidentifikasi berdasarkan DNA sel darah. Hasilnya, seluruh CFU-EC secara konsisten menunjukkan mutasi homozigot dan heterozigot pada gen JAK2, sedangkan hanya 3 ECFC yang termutasi dan seluruhnya berasal dari individu yang sama. Hal ini menunjukkan bahwa CFU-EC berasal dari sel hematopoietik dan tidak dapat berdiferensiasi secara sempurna menjadi sel endotel. Selain itu, CFU-EC tidak dapat menghasilkan koloni sekunder ketika diisolasi dan dikultur pada medium yang sama, sedangkan 32% ECFC dapat menghasilkan koloni sekunder 7 hari setelah kultur, menandakan kemampuan klonogenik (memperbarui diri) dan proliferatif sebagai karakteristik sel punca (termasuk sel progenitor). Lebih lanjut, penelitian ini menunjukkan bahwa re-kultur formasi sel bundar pada bagian sentral sebagai ciri khas CFU-EC mampu berdiferensiasi menjadi CFU-GM, sehingga makin menguatkan bukti bahwa CFU-EC berasal dari turunan sel hematopoietik.
Tabel 1. Karakterisasi CFU-EC dan ECFC berdasarkan Ekspresi Antigen Permukaan 18
dan Identitas Fenotipe lain.18
|
Jenis Karakterisasi |
CFU-EC |
ECFC | |
|
Antigen |
CD31 |
92,31±5,47 (%) |
92,29±1,32 (%) |
|
CD105 |
74,36±6,32 (%) |
96,73±1,79 (%) | |
|
CD144 |
34,80±8,74(%) |
99,15±0,85 (%) | |
|
CD146 |
56,52±10,00 (%) |
94,21±3,71 (%) | |
|
VEGFR2 |
99,19±0,81 (%) |
68,61±11,26 (%) | |
|
vWF |
67,21±12,78 (%) |
97,09±2,05 (%) |
|
UEA-1 |
41,80±11,67 (%) |
100 (%) |
|
Ac-LDL |
73,68±9,05 (%) |
99,75±0,25 (%) |
|
CD14 |
98,53±1,04 (%) |
1,20±0,74 (%) |
|
CD45 |
98,15±1,85 (%) |
0,37±0,37 (%) |
|
CD115 |
94,42±2,52 (%) |
0,28±0,21 (%) |
|
Antigen sel endotel |
Ya |
Ya |
|
Antigen sel hematopoietik |
Ya |
Tidak |
|
Antigen makrofag & fagositosis |
Ya |
Tidak |
|
Klonogenesis sekunder |
CFU-GM |
Sel endotel |
|
Formasi pembuluh darah in vivo |
Tidak |
Ya |
|
Fungsi parakrin |
Ya |
Tidak |
|
Turunan klonal dengan HSC |
Ya |
Tidak |
Identifikasi EPC juga melibatkan morfologi ketika dikultur. Apabila pada medium kultur 2 dimensi, EPC berproliferasi dengan cepat dan membentuk koloni11,16,17,18,25,26, 12
sel endotel dewasa menunjukkan proliferasi minimal dan berbentuk cobblestone.12
Namun ECFC dilaporkan memiliki struktur yang sama dengan sel endotel dewasa saat 18
dikultur (yakni cobblestone).18
Pada akhirnya, suatu tipe sel dapat dikarakterisasi sebagai EPC secara definitif apabila mampu membentuk formasi tabung secara in vitro dan pembuluh darah in vivo.12 Sel endotel ketika dikultur dalam medium 3 dimensi akan mengalami vakuolisasi, pemanjangan, dan ikatan membentuk struktur mirip tabung atau lumen, mirip pembuluh darah.27 Sehingga, teknik kultur pada medium ini dapat mensimulasikan sel yang diujikan secara nyata, mendekati keadaan sebenarnya pada in vivo.28 Walaupun kemampuan membentuk lumen merupakan ciri khas sel endotel yang tidak dimiliki oleh sel lain, beberapa tipe sel dapat membentuk struktur mirip tabung pada sistem kultur 3 dimensi, yang disebut cord-like structures (Gambar 1A & B).28 Struktur cord adalah pemanjangan bentuk sel ke arah eksternal dan membentuk jalinan antara satu dengan lainnya pada medium kultur hingga menyerupai lumen. Salah satu tipe sel yang dapat membentuk struktur ini adalah sel epitel mammae. Namun demikian, struktur cord
masih dapat dibedakan dari formasi lumen yang sesungguhnya oleh sel endotel, karena kompleksitas formasi yang berbeda.12 Dalam proses pembentukan lumen oleh sel
endotel, diperlukan vakuolisasi dan jajaran khusus, sedangkan dalam formasi cord
28
hanya diperlukan pemanjangan dan jalinan sel (Gambar 1C).28
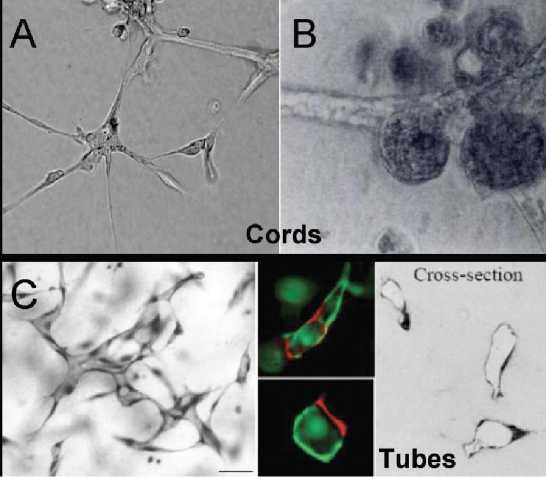
Gambar 1. A) Formasi cord oleh sel endotel, B) formasi cord oleh sel epitel mammae, C) formasi tabung atau lumina oleh sel endotel.12,28
Medium kultur yang digunakan dalam formasi lumen in vitro melibatkan kolagen dan fibrin. Kolagen dianggap penting karena terkandung di dalam matriks ekstraseluler yang terpapar secara konstan pada sel endotel hingga mengekspresikan fenotipe-nya secara optimal in vivo.29 Sedangkan matriks fibrin akan berinteraksi dengan sel endotel vaskuler ketika terjadi perlukaan di daerah lumen yang memicu konversi fibrinogen menjadi fibrin dalam plasma darah yang bersirkulasi, model inflamasi yang sering dijumpai pada saat terjadi iskemia maupun infark.12 Melalui metode ini, dapat dibuktikan bahwa CFU-EC hasil isolasi Asahara et al11 gagal membentuk formasi lumen, sedangkan ECFC mampu membentuk formasi lumen dengan kecepatan yang sama seperti sel endotel dewasa. Selain itu, ECFC mampu melakukan inkorporasi
(penggabungan) dengan sel endotel dewasa hingga membentuk lumen yang tertutup secara sempurna, sedangkan CFU-EC tidak.30 Observasi lanjutan menunjukkan bahwa CFU-EC dapat meningkatkan kemampuan tubulogenesis sel endotel dewasa yang tidak dijumpai pada penggunaan ECFC. Kemampuan EPC membentuk pembuluh darah secara in vivo akan dijelaskan pada subbab berikutnya.
Peran / Fungsi EPC in vivo
EPC berperan dalam Proses Angiogenesis dan Neovaskularisasi
Neovaskularisasi merupakan salah satu strategi reperfusi aliran darah untuk mencegah kematian jaringan akibat iskemia.3,5 EPC yang berasal dari sumsum tulang diketahui mampu bermigrasi menuju daerah iskemia dan mengekspresikan antigen sel endotel, mengindikasikan adanya proses diferensiasi.31 Namun observasi menggunakan mikroskop 3 dimensi konfokal menunjukkan tingkat transdiferensiasi HSC menjadi sel endotel yang relatif rendah.32 Studi lain menunjukkan bahwa injeksi EPC yang telah dimodifikasi secara ex vivo pada hewan coba dengan infark miokardium dapat memperbaiki aliran darah dan kontraktilitas ventrikel secara signifikan.33,34
Isolasi EPC dari sumsum tulang yang telah diberi penanda β-galactosidase (LacZ+) dan diinjeksikan pada mencit dengan aterosklerosis menunjukkan proses diferensiasi EPC menjadi sel endotel dewasa dan terimplantasi secara sempurna pada pembuluh darah dengan tetap mengekspresikan LacZ (Gambar 2).35 Konsisten dengan hal tersebut, beberapa studi klinis yang mentransplantasikan bone marrow-derived stem cells (termasuk EPC di dalamnya) menunjukkan perbaikan reperfusi pada daerah iskemia.36,37 Selain itu, transplantasi EPC yang telah dimodifikasi secara ex vivo pada ekstremitas bawah yang mengalami iskemia (peripheral artery disease) menunjukkan peningkatan
ankle brachial index, mengindikasikan adanya perbaikan aliran darah menuju daerah
iskemia.38
Gambar 2. EPC berperan dalam angiogenesis. Segmen pembuluh darah mencit wildtype diimplantasikan pada xenograf A) ApoE+/+ dan B) ApoE-/-. Segmen diisolasi 4 minggu pasca operasi dan diwarnai menggunakan X-gal. Pembuluh darah mikro berwarna biru, mengindikasikan peranan EPC dalam angiogenesis pada menit B) ApoE-/-.35
CFU-EC dilaporkan memiliki kemampuan vaskulogenesis in vivo sehingga meningkatkan proses neovaskularisasi.39 Namun hal tersebut diduga hanya bersifat parakrin dan diferensiasi menjadi sel endotel tidak dapat dijumpai.22,30 Pada penelitian Yoder et al18, masing-masing CFU-EC dan ECFC dari sumber yang sama dikultur dan ditransplantasikan pada mencit SCID (severe combined immunodeficiency). Pada periode 14 hingga 30 hari, pembuluh darah mencit diekstraksi dan diobservasi menggunakan anti-human CD31. Hasilnya, CFU-EC gagal membentuk pembuluh darah, sedangkan ECFC mampu membentuk lumen pembuluh darah dengan anti-human CD31+ yang menunjukkan bahwa ECFC berasal dari donor (manusia) dan bukan resipien (hewan coba).
Populasi sel lain (CAC) yang mengalami adhesi spontan saat prosedur kultur fase
pertama diujicobakan in vivo dan terbukti memperbaiki neovaskularisasi.40 CAC mengekspresikan CD14 yang berarti merupakan tipe sel monosit/makrofag.12,18 Namun 12
CAC tidak terbukti mampu membentuk formasi lumen dan pembuluh darah.12 Sehingga, peranan CAC pada angiogenesis dan neovaskularisasi diduga kuat bersifat parakrin dengan sekresi kemokin (MCP-1, VEGF, Hematopoietic Growth Factor/HGF, dan G-CSF) yang berfungsi merekrut EPC dari sumsum tulang menuju daerah iskemia.41 Walaupun hanya bersifat parakrin, peranan CAC dalam angiogenesis relatif krusial. Hal ini dibuktikan dengan penurunan arteriogenesis dan Placental Growth Factor (PIGF) saat terjadi deplesi sel mononuklear.
Pada keadaan normal, tingkat inkorporasi EPC pada pembuluh darah relatif rendah. Saat terjadi perlukaan atau iskemia, persentase inkorporasi meningkat hingga 90%.42 Dua studi mengonfirmasikan rerata inkorporasi sel berfenotipe endotel yang berasal dari sumsum tulang (dapat berupa CFU-EC, CAC, atau ECFC) sebesar 34%43, sedangkan studi lain tidak menemukan adanya inkorporasi.44 Hasil yang bervariasi ini dapat dijelaskan oleh variasi model iskemia (intensitas dan tipe jejas) sehingga mempengaruhi hasil akhir. Seperti yang diketahui, iskemia ringan akan menghasilkan proses inkorporasi yang lebih rendah dibandingkan iskemia berat.
EPC berperan dalam Regenerasi Sel Endotel
EPC mampu berdiferensiasi menjadi sel endotel dan melapisi daerah yang terdenudasi. Hal ini dibuktikan dengan implantasi sel CD133+ VEFGR2+ pada permukaan alat pacu ventrikel in vivo.45 Sirkulasi EPC dalam darah juga dilaporkan bermigrasi ke daerah arteri yang terdenudasi pasca intervensi perkutan.46 Hal ini konsisten dengan temuan
studi lain yang menunjukkan implantasi bone marrow-derived stem cells pada arteri yang terdenudasi akibat perlukaan.47 Tetapi implantasi EPC dari kultur sel yang berasal dari sumsum tulang hanya berdiferensiasi menjadi sel endotel dengan intensitas rendah.48 Perbedaan ini dapat kembali dijelaskan dengan adanya subklasifikasi EPC menjadi CFU-EC, CAC, dan ECFC yang tidak dideksripsikan dalam studi yang bersangkutan. Namun secara umum, implantasi EPC menggantikan endotel vaskuler yang terdenudasi berperan penting dalam mempertahankan elastisitas pembuluh darah melalui sekresi NO serta menghambat proliferasi neointima, sehingga memperbaiki prognosis pasien dengan penyakit jantung iskemik.49
Mekanisme Mobilisasi, Homing (Migrasi), dan Diferensiasi EPC
Saat terjadi iskemia maupun jejas vaskuler, terjadi sekresi dan upregulasi vascular endothelial growth factor (VEGF) dan stromal-derived factor-1 (SDF-1) yang dilepaskan menuju sirkulasi dan memodulasi ekspresi matrix metalloproteinase-9 (MMP-9), elastase, dan cathepsin G.5 MMP-9 kemudian akan bergerak menuju sumsum tulang dan memisahkan ikatan antara jaringan ikat (stroma) dengan EPC. MMP-9 terbukti dapat memotong ligan membrane-bound Kit (mKitL) dan memicu pelepasan ligan Kit solubel (sKitL; atau disebut juga stem cell factor/SCF).50 Aktivasi pro-MMP-9 menjadi MMP-9 juga melibatkan aktivitas NO.51 Pada model hewan coba dengan deplesi NO, ekspresi MMP-9 menurun secara signifikan bahkan setelah stimulasi menggunakan VEGF.51 Akibatnya, hewan tersebut tidak responsif terhadap iskemia dan menunjukkan defek pada mobilisasi dan migrasi EPC pada daerah jejas.
EPC yang terlepas dari sumsum tulang akan segera beredar dalam sirkulasi darah dan mengalami aktivasi berbagai reseptor, seperti CXCR4, P-selectin glycoprotein ligand-1
(PGSL-1), dan integrin (α4β1, β2, αvβ3, αvβ5) [Gambar 3].52 Sekresi SDF-1 oleh platelet yang dimodulasi oleh berbagai sitokin proinflamasi hasil sekresi makrofag (MCP-1) dan VEGF akan berikatan dengan chemokine receptor (CXCR4) dan membantu homing (migrasi) EPC menuju daerah jejas.53,54 Setelah berada pada daerah perlukaan, EPC berinteraksi dengan sel endotel yang terluka seperti interaksi makrofag dengan endotel vaskuler. Endotel yang luka akan meng-upregulasi ekspresi Intracellular Adhesion Molecule (ICAM-1), P-selectin, dan E-selectin yang berfungsi dalam proses adhesi EPC melalui ikatan dengan integrin dan P-Selectin Glycoprotein Ligand (PSGL-1) yang diekspresikan pada permukaan EPC.55 Aktivasi EphB4 dilaporkan dapat meningkatkan ekspresi PSGL-1 di permukaan EPC56, sedangkan sitokin high-mobility group box 1
(HMGB-1) yang dilepaskan oleh endotel nekrosis dapat mengaktivasi integrin β1 dan β2
57
pada permukaan EPC.57
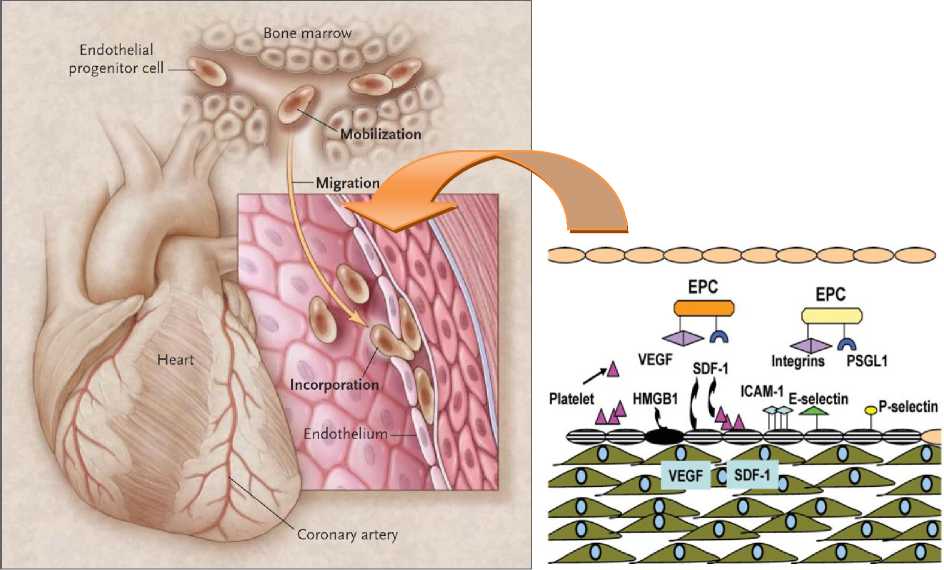
Gambar 3. Mekanisme Mobilisasi, Migrasi, dan Diferensiasi EPC. EPC dimobilisasi dari sumsum tulang dan berdiferensiasi menjadi sel endotel pada daerah jejas vaskuler. (Kanan) ekspresi berbagai antigen permukaan, reseptor, dan sitokin.1,52
Saat dalam perjalanan menuju daerah jejas, EPC berdiferensiasi menjadi sel endotel yang dioptimasi oleh beragam sitokin (termasuk VEGF dan SDF-1) serta tekanan mekanik aliran darah (shear stress), sehingga EPC dapat mengekspresikan fenotipe sel endotel dewasa dan gen-gen spesifik endotel vaskuler.5,52 Setelah proses diferensiasi selesai dan EPC telah menjadi sel endotel dewasa, sel-sel ini akan bersifat invasif dengan melakukan adhesi (seperti dijelaskan pada kondisi kultur in vitro) pada lumen vaskuler. Proses ini tercapai dengan bantuan protease, cathepsin, dan MMP-2 yang bersifat destruktif terhadap komponen sel yang telah rusak sehingga membantu implantasi endotel baru yang menggantikan sel-sel endotel yang terluka dan mati, sehingga meningkatkan sekresi NO yang berperan dalam proses rekrutmen, neovaskularisasi, dan reperfusi area iskemik.5
KESIMPULAN DAN SARAN
EPC berperan penting dalam proses neovaskularisasi dan angiogenesis demi mempertahankan integritas vaskuler saat terjadi iskemia maupun inflamasi akibat jejas seluler. Kapasitas intrinsik EPC telah dibuktikan dalam berbagai penelitian in vitro dan telah dibuktikan dalam studi klinis. Namun beberapa hal perlu diklarifikasi lebih lanjut, diantaranya adalah definisi EPC secara spesifik. Saat ini telah terdapat tiga subtipe EPC berbeda (CFU-EC, CAC, ECFC) yang masing-masing memiliki karakteristik fenotipe, klonogenesis, prosedur kultur, serta fungsionalitas yang berbeda pula. Oleh karena itu, perlu dibentuk klasifikasi baru yang menetapkan definisi EPC, terutama berdasarkan
kemampuan membentuk formasi tabung/lumen in vitro dan pembuluh darah secara in
vivo serta efek parakrin yang masing-masing hanya dimiliki oleh ECFC dan CFU-EC, CAC, secara berurutan.
Melalui kategorisasi ini akan memperjelas masing-masing subtipe EPC dan mempermudah pengembangan strategi terapi optimal per subtipe yang disesuaikan berdasarkan kapasitas utamanya. Misal, apabila ECFC telah divalidasi menjadi subtipe yang dapat membentuk pembuluh darah dan tidak bersifat parakrin, maka segala hal yang berkaitan dengan ekspansi/modifikasi kultur in vitro akan difokuskan pada kemampuan ECFC dalam membentuk formasi lumen. Demikian juga sebaliknya. Apabila hal ini terealisasi, maka akan muncul pertanyaan baru mengenai manakah strategi terapi yang paling efektif dan efisien dalam menginduksi neovaskularisasi pada manusia? Apakah terapi transplantasi ECFC atau terapi angiogenik berbasis sitokin untuk memodulasi fungsi EPC internal?
Apabila aktivitas biologi seluler EPC beserta karakteristik fenotipe, morfologi, dan fungsionalitasnya telah dapat dipetakan secara jelas, maka aplikasi EPC dalam terapi klinis pada penderita penyakit jantung iskemik tinggal selangkah lagi.
DAFTAR PUSTAKA
-
1. Rosenzweig A. Endothelial progenitor cells. N Engl J Med 2003; 348(17): 581-2.
-
2. Shantsila E, Watson T, Lip YH. Endothelial progenitor cells in cardiovascular disorders. J Am Coll Cardiol 2007; 49; 741-52.
-
3. Sasaki K, Heeschen C, Mildner-Rihm C, Urbich C, Ihling C, Technau-Ihling et al. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat Med 2003; 9: 1370-6.
-
4. Wollert KC, Drexler H. Clinical applications of stem cells for the heart. Circ Res 2005; 96: 151-63.
-
5. Urbich C, Dimmeler S. Endothelial progenitor cells: characterization and role in vascular biology. Circ Res 2004; 95: 343-53.
-
6. Dimmeler S, Burchfield J, Zeiher AM. Cell-based therapy of myocardial infarction. Arterioscler Thromb Vasc Biol 2008; 18: 33-7.
-
7. Papayannopoulou T. Current mechanistic scenarios in hematopoietic stem/progenitor cell mobilization. Blood 2004; 103: 1580–5.
-
8. Heissig B, Hattori K, Dias S, Friedrich M, Ferris B, Hackett NR, et al. Recruitment of stem and progenitor cells from the bone marrow niche requires MMP-9 mediated release of kit-ligand. Cell 2002; 109: 625–37.
-
9. Lee SH, Wolf PL, Escudero R, Deutsch R, Jamieson SW, Thistlethwaite PA. Early expression of angiogenesis factors in acute myocardial ischemia and infarction. N Engl J Med 2000; 342: 626–33.
-
10. Pillarisetti K, Gupta SK. Cloning and relative expression analysis of rat stromal cell derived factor-1 (SDF-1)1: SDF-1 alpha mRNA is selectively induced in rat model of myocardial infarction. Inflammation 2001; 25: 293–300.
-
11. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997; 275; 964-7.
-
12. Hirschi K, Ingram DA, Yoder MC. Assessing identity, phenotype, and fate of endothelial progenitor cells. Arterioscler Thromb Vasc Biol 2008; 28: 1584-95.
-
13. Badorff C, Brandes RP, Popp R, Rupp S, Urbich C, Aicher A, et al. Transdifferentiation of blood-derived human adult endothelial progenitor cells into functionally active cardiomyocytes. Circulation 2003; 107: 1024 –32.
-
14. Woywodt A, Blann AD, Kirsch T, Erdbruegger U, Banzet N, Haubitz M, et al. Isolation and enumeration of circulating endothelial cells by immunomagnetic isolation: proposal of a definition and a consensus protocol. J Thromb Haemost 2006; 4: 671– 7.
-
15. Shi Q, Rafii S, Wu MH, Wijelath ES, Yu C, Ishida A, et al. Evidence for circulating bone marrow-derived endothelial cells. Blood 1998; 92: 362–7.
-
16. Ito H, Rovira II, Bloom ML, Takeda K, Ferrans VJ, Quyyumi AA, et al. Endothelial progenitor cells as putative targets for angiostatin. Cancer Res 1999; 59: 5875–7.
-
17. Hill JM, Zalos G, Halcox JP, Schenke WH, Waclawiw MA, Quyyumi AA. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N Engl J Med 2003;348:593– 600.
-
18. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, et al. Redefining endothelial progenitor cells via clonal analysis and hematopoietic stem/progenitor cell principals. Blood 2007; 109: 1801–9.
-
19. Vasa M, Fichtlscherer S, Adler K, Aicher A, Martin H, Zeiher AM, et al. Increase in circulating endothelial progenitor cells by statin therapy in patients with stable coronary artery disease. Circulation 2001; 103: 2885–90.
-
20. Vasa M, Fichtlscherer S, Aicher A, Adler K, Urbich C, Martin H, et al. Number and migratory activity of circulating endothelial progenitor cells inversely correlate with risk factors for coronary artery disease. Circ Res; 2001; 89: E1–7.
-
21. Hassan N, Campbell D, Douglas S. Purification of human monocytes on gelatin-coated surfaces. J Immunol Methods 1986; 95: 273–76.
-
22. Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, et al. Ischemia-and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med 1999; 5: 434–8.
-
23. Fujiyama S, Amano K, Uehira K, Yoshida M, Nishiwaki Y, Nozawa Y, et al. Bone marrow monocyte lineage cells adhere on injured endothelium in a monocyte chemoattractant protein-1-dependent manner and accelerate reendothelialization as endothelial progenitor cells. Circ Res 2003; 93: 980–9.
-
24. Zhang SJ, Zhang H, Wei YJ, et al. Adult endothelial progenitor cells from human peripheral blood maintain monocyte/macrophage function throghout in vitro culture. Cell Res 2006; 16: 577-84.
-
25. Lin Y, Weisdorf DJ, Solovey A, Hebbel RP. Origins of circulating endothelial cells and endothelial outgrowth from blood. J Clin Invest 2000; 105: 71–7.
-
26. Ingram DA, Mead LE, Tanaka H, Meade V, Fenoglio A, Mortell K, et al. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004; 104: 2752–60.
-
27. Madri JA, Pratt BM. Endothelial cell-matrix interactions: in vitro models of angiogenesis. J Histochemistry Cytochemistry 1986; 34: 85–91.
-
28. Davis GE, Wonshill K, Stratman AN. Mechanisms controlling human endothelial lumen formation and tube assembly in three-dimensional extracellular matrices. Birth Defects Res 2007; 81: 270 –85.
-
29. Delvos U, Gajdusek C, Sage H, Harker LA, Schwartz SM. Interactions of vascular wall cells with collagen gels. Lab Investigation 1982; 46: 61–72.
-
30. Sieveking DP, Buckle A, Celermajer DS, Ng MKC. Strikingly different angiogenic properties of endothelial progenitor cell subpopulations. J Am Coll Cardiol 2008; 51: 660–8.
-
31. Xu Q, Zhang Z, Davison F, Hu Y. Circulating progenitor cells regenerate endothelium of vein graft atherosclerosis, which is diminished in apoE deficient mice. Circ Res 2003; 93: e76–86.
-
32. Purhonen S, Palm J, Rossi D, Kaskenpaa N, Rajantie I, Hla-Herttuala S, et al. Bone marrow-derived circulating endothelial precursors do not contribute to vascular endothelium and are not needed for tumor growth. Proc Natl Acad Sci U S A 2008; 105: 6620–5.
-
33. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, et al. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc Natl Acad Sci U S A 2000; 97: 3422–7.
-
34. Kawamoto A, Gwon HC, Iwaguro H, Yamaguchi JI, Uchida S, Masuda H, et al. Therapeutic potential of ex vivo expanded endothelial progenitor cells for myocardial ischemia. Circulation 2001; 103: 634–7.
-
35. Hu Y, Davison F, Zhang ZG, Xu Q. Endothelial replacement and angiogenesis in arteriosclerotic lesions of allografts are contributed by circulating progenitor cells. Circulation 2003; 108: 3122–7.
-
36. Schächinger V, Erbs S, Elsässer A, et al. Intracoronary bone marrow–derived progenitor cells in acute myocardial infarction. N Engl J Med 2006; 355: 1210-21.
-
37. Assmus B, Schächinger V, Teupe C, et al. Transplantation of Progenitor Cells and Regeneration Enhancement in Acute Myocardial Infarction (TOPCARE-AMI). Circulation 2002; 106: 3009-17.
-
38. Tateishi-Yuyama E, Matsubara H, Murohara T, Ikeda U, Shintani S, et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: a pilot study and a randomised controlled trial. Lancet 2002; 360: 427–35.
-
39. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, et al. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol 2004; 24: 288–93.
-
40. Urbich C, Heeschen C, Aicher A, Dernbach E, Zeiher AM, Dimmeler S. Relevance of monocytic features for neovascularization capacity of circulating endothelial progenitor cells. Circulation 2003;108: 2511–6.
-
41. van Royen N, Hoefer I, Buschmann I, Kostin S, Voskuil M, Bode C, et al. Effects of local MCP-1 protein therapy on the development of the collateral circulation and atherosclerosis in Watanabe hyperlipidemic rabbits. Cardiovasc Res 2003; 57: 178– 85.
-
42. Jackson KA, Majka SM, Wang H, Pocius J, Hartley CJ, Majesky MW, et al. Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells. J Clin Invest 2001; 107: 1395–402.
-
43. Zhang ZG, Zhang L, Jiang Q, Chopp M. Bone marrow-derived endothelial progenitor cells participate in cerebral neovascularization after focal cerebral ischemia in the adult mouse. Circ Res 2002; 90: 284–8.
-
44. Hess DC, Hill WD, Martin-Studdard A, Carroll J, Brailer J, Carothers J. Bone marrow as a source of endothelial cells and NeuN-expressing cells After stroke. Stroke 2002; 33: 1362–8.
-
45. Peichev M, Naiyer AJ, Pereira D, Zhu Z, Lane WJ, Williams M, et al. Expression of VEGFR-2 and AC133 by circulating human CD34(_) cells identifies a population of functional endothelial precursors. Blood 2000;95:952–8.
-
46. Walter DH, Rittig K, Bahlmann FH, Kirchmair R, Silver M, Murayama T, et al. Statin therapy accelerates reendothelialization: a novel effect involving mobilization and incorporation of bone marrow-derived endothelial progenitor cells. Circulation 2002; 105: 3017–24.
-
47. Werner N, Junk S, Laufs U, Link A, Walenta K, Bohm M, et al. Intravenous transfusion of endothelial progenitor cells reduces neointima formation after vascular injury. Circ Res 2003; 93: e17–24.
-
48. Hillebrands JL, Klatter FA, van Dijk WD, Rozing J. Bone marrow does not contribute substantially to endothelial-cell replacement in transplant arteriosclerosis. Nat Med 2002; 8: 194–5.
-
49. Fujiyama S, Amano K, Uehira K, Yoshida M, Nishiwaki Y, Nozawa Y, et al. Bone marrow monocyte lineage cells adhere on injured endothelium in a monocyte chemoattractant protein-1-dependent manner and accelerate reendothelialization as endothelial progenitor cells. Circ Res 2003; 93:980–9.
-
50. Heissig B, Hattori K, Dias S, Friedrich M, Ferris B, Hackett NR, et al. Recruitment of stem and progenitor cells from the bone marrow niche requires MMP-9 mediated release of kit-ligand. Cell 2002; 109: 625–37.
-
51. Aicher A, Heeschen C, Mildner-Rihm C, Urbich C, Ihling C, Technau-Ihling K, et al. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat Med 2003; 9: 1370–6.
-
52. Zampetaki A, Kirton JP, Xu Q. Vascular repair by endothelial cells. Cardiovas Res 2008; 78; 413-21.
-
53. Massberg S, Konrad I, Schurzinger K, Lorenz M, Schneider S, Zohlnhoefer D, et al. Platelets secrete stromal cell-derived factor-1 alpha and recruit bone marrow-derived progenitor cells to arterial thrombi in vivo. J Exp Med 2006; 203: 1221–33.
-
54. Hiasa K, Ishibashi M, Ohtani K, Inoue S, Zhao Q, Kitamoto S, et al. Gene transfer of stromal cell-derived factor-1alpha enhances ischemic vasculogenesis and angiogenesis via vascular endothelial growth factor/endothelial nitric oxide synthase-related pathway: next-generation chemokine therapy for therapeutic neovascularization. Circulation 2004; 109: 2454–61.
-
55. Vajkoczy P, Blum S, Lamparter M, Mailhammer R, Erber R, Engelhardt B, et al. Multistep nature of microvascular recruitment of ex vivo-expanded embryonic endothelial progenitor cells during tumor angiogenesis. J Exp Med 2003; 197: 1755–65.
-
56. Foubert P, Silvestre JS, Souttou B, Barateau V, Martin C, Ebrahimian TG et al. PSGL-1-mediated activation of EphB4 increases the proangiogenic potential of endothelial progenitor cells. J Clin Invest 2007;117: 1527–37
-
57. Chavakis E, Hain A, Vinci M, Carmona G, Bianchi ME, Vajkoczy P et al. High-mobility group box 1 activates integrin-dependent homing of endothelial progenitor cells. Circ Res 2007; 100: 204–12.
22
Discussion and feedback