EKSPLORASI SPASIAL CENDAWAN TANAH PADA SEKITAR RHIZOSFER TANAMAN JAMBU METE (Anacardium occidentale L.) DI KARANGASEM DAN BULELENG-BALI
on
JURNAL SIMBIOSIS I (2): 85- 101
Jurusan Biologi FMIPA Universitas Udayana
ISSN : 2337-7224
September 2013
EKSPLORASI SPASIAL CENDAWAN TANAH PADA SEKITAR RHIZOSFER TANAMAN JAMBU METE (Anacardium occidentale L.)
DI KARANGASEM DAN BULELENG-BALI
(SPACIAL EXPLORATION OF FUNGI IN RHIZOSPHERE A CASHEW PLANTATIONS (Anacardium occidentale L.) AT KARANGASEM AND BULELENG-BALI)
Ni Luh Desy Wulandari1), Meitini W. Proborini2), I Ketut Sundra2) Jurusan Biologi, FMIPA, Universitas Udayana
*) Email: Niluhdesy_wulandari@yahoo.com
INTISARI
Penelitian telah dilaksanakan yang bertujuan untuk mengetahui total koloni, keanekaragaman, dan tingkat penyebaran cendawan tanah yang terdapat pada rizosfer perkebunan jambu mete (Anacardium occidentale L.) di Sukadana Karangasem dan Sendang Buleleng. Penelitian ini dilaksanakan selama empat bulan (Oktober 2012 – Januari 2013). Sampel dari dua lokasi berbeda di Sukadana Karangasem dan Sendang Buleleng, masing-masing lokasi diambil dari 5 titik yang berbeda. Hasil data yang diidentifikasi jenis cendawan secara deskripsi. Data total koloni yang dikuantifikasi dengan rumus yang telah ditetapkan. Hasil penelitian teridentifikasi 11 jenis cendawan pada kedua lokasi yang berbeda yaitu : Aspergilus flavus, A. niger, A. parasiticus, Botrytis cinera, Cladosporium sphaeospermum, Fusarium sporotrichioides, P. brevicompactum, P. citrinum, P. chrysogenum, Rhizopus stolonifer, dan Synchephalastrum racemosum. Hasil perhitungan total koloni cendawan yang didapatkan di Sukadana Karangasem dengan total rata-rata adalah 33,6 x 103 – 35 x103 CFU/g, sedangkan lokasi Sendang Buleleng 22,6 x 103 – 37,6 x 103 CFU/g. Tingkat penyebaran cendawan di Karangasem adalah tergolong seragam (0,85) dan di Buleleng tergolong mengelompok (1,03).
Kata kunci: Rhizosfer mete, keanekaragaman cendawan, Karangasem, Buleleng-Bali
ABSTRACT
Research have been carried out to calculate total colony of fungi, determine fungi species, and the rate spread of the fungi in rhizosphere a cashew plantations (Anacardium occidentale L.) at Sukadana Karangasem and Sendang Buleleng for 4 months (October 2012 - January 2013). Samples were taken from two different areas in which five point were sampled. Results data were analyzed qualitatively by identifying the species of fungi that is a description. The quantitative data was taken to calculate total colony of fungi. The research found 11 species of fungi taken from two different locations, namely: Aspergilus flavus, A. niger, A. parasiticus, Botrytis cinera, Cladosporium sphaeospermum, Fusarium sporotrichioides, P. brevicompactum, P. citrinum, P. chrysogenum, Rhizopus stolonifer, and Synchephalastrum racemosum. Total fungal colony in Sukadana Karangasem was : 33.6 x 103 - 35 x 103 CFU /g, in Sendang Buleleng 22.6 x 103 – 37.6 x 103 CFU/g. Abundance of fungi in Sukadana Karangasem is relatively uniform (0.85) and in Sendang Buleleng belong to cluster (1,03).
Keywords: Cashew rhizosphere, abudance of fungal species, Karangasem, Buleleng-Bali
PENDAHULUAN
Cendawan tanah banyak memiliki peranan penting dalam dekomposisi tanah (Gandjar dkk., 2006). Kelompok cendawan anggota ordo Peronosporales, Mucorales, dan beberapa dari kelas Ascomycetes atau Deuteromycetes yang bersifat saprofit ditanah. Menurut Dharmaputra dkk., 1989 Cendawan di rizosfer berbentuk miselium, spora (konidia), dan khlamidospora. Miselium berfungsi untuk melakukan aktifitas metabolisme. Populasi cendawan di rizosfer biasanya lebih banyak dan beragam dibandingkan pada tanah bukan rizosfer (Lynch 1990; Carlile et al.2001). Menurut Novriani dan Madjid (2009) terjadinya interaksi antara mikroorganisme dengan tanaman pada rizosfer dapat bersifat simbiosa mutualisme, parasitisme atau kompetisi. Interaksi tersebut dapat mempengaruhi pertumbuhan tanaman, baik interaksi yang menguntungkan atau merugikan tanaman. Interaksi antara mikroorganisme dengan tanaman (baik yang menguntungkan maupun merugikan) akan berkompetisi pada lingkungan tersebut yang disertai oleh pengaruh faktor eksternal dan faktor internal. Kedua faktor tersebut akan memberikan reaksi yang berbeda terhadap pertumbuhan tanaman.
Menurut Darmawijaya (1990) perbedaan tekstur tanah dapat berpengaruh terhadap jumlah populasi dan keanekaragaman mikroorganisme disekitarnya (cendawan dan bakteri) maupun makroorganisme (fauna tanah). Perbedaan lokasi tersebut sangat berkaitan dengan struktur tanah sehingga adanya perbedaan pada kedua lokasi. Sampel tanah yang diambil dari Buleleng mempunyai tekstur tanah lempung berdebu. Menurut Hanafiah dkk., (2005) tanah-tanah lempung berdebu memiliki sifat tanah seperti berikut : tekstur licin, membentuk butiran/menggumpal keras, mengkilat, dan memiliki ruang pori yang berukuran kecil, sehingga menyebabkan tanah-tanah tersebut pada waktu musim hujan sangat liat dan pada waktu musim kemarau sangat keras. Sampel tanah di Karangasem dengan kondisi tanah lempung berpasir. Menurut Hanafiah dkk., (2005) tanah-tanah lempung berpasir memiliki sifat tanah seperti berikut: Tekstur kasar, membentuk butiran yang agak keras tetapi mudah hancur, tidak melekat, memiliki ruang pori yang berukuran besar.
Berdasarkan latar belakang yang sudah dipaparkan di atas, penulis ingin meneliti jenis-jenis cendawan apa saja
yang terdapat pada rizosfer perkebunan tanaman jambu mete (Anacardium occidentale L.) pada kedua lokasi tersebut. Tempat dan Waktu Penelitian
Penelitian dilaksanakan di Laboratorium Taksonomi Tumbuhan (Mikologi). Analisis tanah dilaksanakan di Laboratorium Ilmu Tanah Fakultas Pertanian Universitas Udayana. Penelitian ini dilaksanakan selama empat bulan (Oktober 2012 – Januari 2013).
Teknik Pengambilan Sampel
Pengambilan sampel dilakukan dengan metode jelajah/eksplorasi yaitu menentukan titik-titik pengambilan sampel (secara acak dengan cara membuat pengundian yang ditentukan pada lima titik pengambilan sampel), untuk menghubungkan titik satu dengan titik yang lainnya diberikan jarak ± 200 meter. Sampel tanah diambil pada rizosfer/area di sekeliling akar tanaman mete dengan kedalaman 10-20 cm dengan menggunakan skop. Sampel tanah dari lima tanaman mete dicampur (komposit) dan diambil ± 100 gram, kemudian tiap-tiap titik pengambilan sampel yang lainya (sampai titik ke lima) dilakukan dengan cara yang sama.
Isolasi Cendawan dan Perhitungan Total Koloni Cendawan
Isolasi cendawan dilakukan dengan metode pour plate (Pelczar dan Chan, 2006). Isolasi cendawan tanah dilakukan pada pengenceran 10-3. Pada pengenceran 10-1 botol yang telah terisi air steril sebanyak 90 ml ditambah sampel tanah sebanyak 10 gram kemudian dikocok hingga homogen, pengenceran 10-2 tabung reaksi yang telah terisi air steril sebanyak 9 ml kemudian ditambahkan 1 ml suspensi sampel tanah dari pengenceran sebelumnya kemudian di vortex langkah ini dilakukan sampai pengenceran 10-5 dari pengenceran tersebut masing-masing dituang dalam cawan petri steril, kemudian media PDA dituang ke dalam petri pada suhu 400 C dan cawan petri diputar hingga homogen, selanjutnya dilakukan inkubasi di dalam enkas (tempat inkubasi cendawan) selama 2 – 3 hari pada suhu ruang 25 – 270C. Hifa yang tumbuh kemudian diamati dan dipindahkan ke media baru (reisolasi), selanjutnya diidentifikasi, pada penelitian ini dilakukannya tiga kali ulangan (Proborini, 2002).
Total koloni cendawan yang tumbuh pada media PDA, akan tampak warna awal yaitu berwarna putih, kemudian diamati, dipilih yang dianggap
memiliki ciri-ciri sperti cendawan. Dihitung total koloni dengan
menggunakan rumus. Rumus yang digunakan untuk menghitung total koloni cendawan yaitu:
Jumlah cendawan = Jumlah koloni per cawan x1 Faktor_Pengenceran
Dengan satuan CFU (Colony Forming Units) (Kawuri dkk, 2007).
Penyebaran Jenis Cendawan
Untuk mengetahui penyebaran jenis-jenis cendawan di rizosfer perkebunan jambu mete pada kedua lokasi (Sukadana Karangasem dan Sendang Buleleng) dapat digunakan seperti rumus pola penyebaran yang ada dibawah ini :
Rumus :
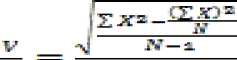
Keterangan : V = Varian M = Mean/rata-rata individu N = Jumlah spesies
X = Jumlah individu masing-masing spesies (Odum, 1993).
Hasil dan Pembahasan
Hasil Total Koloni Cendawan
Hasil perhitungan total koloni di rizosfer perkebunan jambu mete pada lokasi Karangasem adalah antara 33,6 x 103 - 35 x 103 CFU/g, sedangkan lokasi Buleleng 22,6 x 103 - 37,6 103 CFU/g. Data selengkapnya ditampilkan pada Tabel 3 di bawah ini :
Tabel 1.Total rata-rata koloni cendawan pada semua titikpada kedua lokasi (Karangasem dan Buleleng)
Lokasi Sampel/Total Rata-rata Koloni
|
Karangasem KM1 KM2 KM3 KM4 KM5 |
Total Koloni Buleleng |
Total Koloni (CFU/g) 35,3 x 103 22,6 x 103 34,3 x 103 36 x 103 37,6 x 103 | |
|
(CFU/g) 35 x 103 33,3 x 103 34 x 103 32,6 x 103 34,3 x 103 |
BL1 BL2 BL3 BL4 BL5 | ||
|
Keterangan : | |||
|
KM1 = Karangasem titik 1 |
BL1 = Bulelelng titik 1 | ||
|
KM2 = Karangasem titik 2 |
BL2 = Bulelelng titik 2 | ||
|
KM3 = Karangasem titik 3 |
BL3 = Bulelelng titik 3 | ||
|
KM4 = Karangasem titik 4 |
BL4 = Bulelelng titik 4 | ||
|
KM 5 = Karangasem titik 5 |
BL5 = Bulelelng titik 5 | ||
Hasil penelitian menunjukan data tertinggi dan terrendah rata-rata total koloni cendawan yang tumbuh di cawan petri pada masing-masing lokasi Karangasem dan Buleleng dengan menggunakan pengenceran 103. Total koloni cendawan tertinggi terdapat pada KM5 (lokasi Karangasem titk 5) sebesar 34,3 x 103 CFU/g dan BL5 (lokasi Buleleng titk 5) sebesar 37,6 x 103 CFU/g. Total cendawan terendah terdapat pada KM4 (lokasi Karangasem titik 4) yaitu 32,6 x 103 CFU/g dan BL2 (lokasi Buleleng titik 2) yaitu 22,6 x 103 CFU/g (Tabel 1). Tinggi dan rendahnya total koloni cendawan yang didapat pada lokasi Karangasem dan Buleleng, disebabkan karena tidak meratanya penyebaran spora cendawan pada saat pengambilan sampel tanah dimasing-masing titik tersebut. Hasil pengamatan lapangan dan hasil analisis tanah yang telah diujikan dapat mempengaruhi total koloni cendawan yang didapatkan pada kedua lokasi. Hal ini karena faktor lingkungan dan ketersediaan unsur hara pada masing-masing titik yang berperan terhadap pertumbuhan cendawan tersebut.
Menurut Tarigan (1988) pertumbuhan mikroba dipengaruhi oleh lingkungan, baik faktor biotik maupun abiotik. Salah satu faktor lingkungan yang berpengaruh terhadap pertumbuhan mikroorganisme (khususnya cendawan tanah) adalah ketersediaan nutrisi yang dapat menunjang kehidupannya. Menurut Purwaningsih (2005) kesuburan tanah dapat diprediksi dari jumlah populasi mikroba yang hidup di dalamnya. Tingginya jumlah mikroba merupakan pertanda tingginya tingkat kesuburan tanah, karena mikroba berfungsi sebagai perombak senyawa organik menjadi nutrien yang tersedia bagi tanaman, di dalam tanah terkandung cukup bahan organik dan senyawa lainnya untuk pertumbuhan mikroba. Kelembaban tanah berpengaruh pada aerasi, suhu dan reaksi di dalam tanah.
Hasil Identifikasi Cendawan
Berdasarkan hasil identifikasi cendawan yang ditemukan pada lokasi di Karangasem 9 spesies dan di Buleleng 9 spesies, data selengkapnya dapat ditampilkan pada table 2 di bawah ini:
Tabel 2.Jenis dan jumlah cendawan yang ditemukan pada kedua lokasi (Karangasem dan Buleleng)
|
No |
Jenis Cendawan |
Lokasi | |||||||||||
|
KM |
Total |
BL |
Total | ||||||||||
|
1 |
2 |
3 |
4 |
5 |
1 |
2 |
3 |
4 |
5 | ||||
|
1. |
Aspergilus flavus |
2 |
2 |
9 |
- |
7 |
20 |
- |
- |
- |
- |
- |
0 |
|
2. |
Aspergilus niger |
15 |
13 |
11 |
9 |
15 |
65 |
15 |
10 |
14 |
15 |
13 |
68 |
|
3. |
Aspergilus parasiticus |
- |
- |
- |
- |
- |
0 |
. |
2 |
- |
- |
1 |
3 |
|
4. |
Botrytis cinera |
8 |
4 |
10 |
6 |
- |
31 |
5 |
1 |
- |
- |
3 |
11 |
|
5. |
Cladosporium Sphaeospermum |
1 |
- |
- |
2 |
3 |
6 |
2 |
3 |
3 |
- |
- |
8 |
|
6. |
Fusarium Sporotrichioides |
- |
- |
- |
- - |
- |
0 |
- |
- |
1 |
1 |
- |
2 |
|
7. |
Penicilium brevicompactum |
1 |
5 |
- |
8 |
3 |
17 |
5 |
4 |
6 |
8 |
9 |
32 |
|
8. |
Penicilium Citrinum |
6 |
9 |
3 |
7 |
4 |
29 |
- |
- |
- |
- |
- |
0 |
|
9. |
Penicilium Chrysogenum |
- |
- |
2 |
- |
3 |
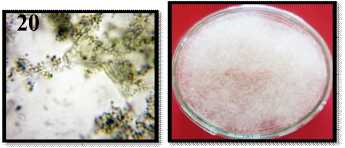
5 |
2 |
- |
4 |
8 |
8 |
22 |
|
10 |
Rhizopus Stolonifer |
11 |
3 |
- |
- |
_ |
15 |
13 |
2 |
10 |
11 |
6 |
42 |
|
11. |
Synchephalastrum racemosum |
2 |
2 |
2 |
- |
2 |
8 |
- |
2 |
4 |
- |
- |
6 |
Cendawan yang ditemukan pada kedua lokasi (Sukadana Karangasem dan Sendang Buleleng) spesies yang diperoleh adalah sebelas 11. Lokasi di Sukadana Karangasem Sembilan 9 spesies dan lokasi Sendang Buleleng Sembilan 9 spesies, tetapi yang dapat membedakan kedua lokasi tersebut adalah spesies dari cendawannya (lokasi Karangasem terdapat Aspergilus flavus dan Penicilium citrinum, sedangkan di Buleleng Aspergilus parasiticus dan Fusarium sporotrichioides). Berdasarkan hasil penelitian pada Tabel 2 menunjukkan, bahwa terjadi keanekaragaman jumlah dan jenis spesies di kedua lokasi (Karangasem dan Buleleng). Keanekaragaman jenis dan jumlah cendawan dipengaruhi oleh faktor lingkungan di kawasan perkebunan jambu mete pada kedua lokasi tersebut. Faktor lingkungan yang mempengaruhi tersebut meliputi analisis tanah yang telah diujikan seperti : ketersediaan unsur hara (C, N, P,
K), tekstur tanah, pH tanah kelembaban, dan suhu menunjukan hasil yang berbeda pada kedua lokasi tersebut.
Berdasarkan hasil analisis tanah pada kedua lokasi, ketersediaan unsur hara (C, N, P, dan K) dapat pula mempengaruhi keberadaan mikroorganisme (cendawan tanah). Hal ini karena beberapa mikroorganisme memiliki peranan penting di dalam tanah untuk penyediaan maupun penyerapan unsur hara bagi tanaman. Tiga unsur hara penting bagi tanaman, yaitu Nitrogen (N), Fosfor (P), dan Kalium (K) seluruhnya melibatkan aktivitas mikroba.
Menurut Wahyuaskari (2010) unsur hara N tersedia melimpah di udara ± 74%, tetapi N di udara tidak dapat langsung dimanfaatkan tanaman. Unsur N harus ditambat atau difiksasi oleh mikroba dan diubah bentuknya menjadi ketertersediaan bagi tanaman, salah satunya amoniak (NH3) menjadi nitrat (HNO3). Mikroba penambat N ada yang
bersimbiosis (Rhizobium sp.) dan ada pula nonsimbiosis (hidup bebas) (Pseudomonas sp.). Mikroba tanah lain yang berperan di dalam penyediaan unsur hara adalah mikroba pelarut fosfat (P) (Aspergilus sp. dan Penicilium sp.) dan kalium (K). Mikroba yang berkemampuan tinggi melarutkan P, umumnya juga berkemampuan tinggi dalam melarutkan K. Kelompok mikroba lain yang juga berperan dalam penyerapan unsur hara.
Menurut Jeger (2001) populasi mikrorganisme baik yang bersifat antagonis, patogen, maupun saprofit dapat menambah keanekaragaman spesies di dalam tanah. Menurut Barker and Weeks (1991) selain faktor-faktor tersebut perbedaan lokasi dan rizosfer menyebabkan perbedaan keanekaragaman spesies dan populasi cendawan. Tekstur tanah yang didominasi oleh fraksi lempung (clay) biasanya mengandung jumlah mikroorganismenya lebih sedikit dibandingkan kondisi tanah dengan tekstur berpasir.
Ditemukan pada kedua lokasi penelitian yaitu genus Aspergilus diantaranya A. niger, A. flavus, dan A. parasiticus. Berdasarkan hasil penelitian pada Tabel 2 cendawan Aspergilus flavus hanya ditemukan pada lokasi Karangasem yaitu dengan total (20 koloni), sedangkan
pada lokasi Buleleng tidak ditemukan cendawan Aspergilus flavus, tetapi pada lokasi Sendang Buleleng ditemukan cendawan Aspergilus parasiticus dengan jumlah (3 koloni). Hal ini karena A. flavus lebih mampu berkompetisi pada lingkungan yang lebih ekstrim (berdasarkan hasil analisis tanah) yaitu di Sukadana Karangasem dibandingkan di Sendang Buleleng, sebaliknya cendawan Aspergilus parasiticus yang lebih mampu berkompetisi di Sendang Buleleng.
Menurut Scheidegger dan Payne (2003) Aspergillus flavus merupakan cendawan saprofit di tanah, pada umumnya memiliki peranan penting sebagai pendaur ulang nutrisi yang terdapat dalam sisa-sisa tumbuhan maupun binatang. Menurut Hedayati et al. (2007) Aspergillus flavus tersebar luas, karena produksi konidianya yang dapat tersebar melalui udara dan melalui serangga, selain itu pertumbuhannya dipengaruhi oleh kadar air, oksigen, dan ketersediaan unsur hara. Menurut Rukmana dkk., (1997) cendawan Aspergillus parasiticus merupakan cendawan yang dapat merusak bahkan merugikan tanaman. Menurut Abbas (2005) cendawan yang termasuk dalam genus Aspergillus sangat dikenal karena peranannya sebagai patogen pada tanaman dan kemampuannya untuk
menghasilkan aflatoksin pada tanaman yang terinfeksi.
Cendawan Aspergilus niger di temukan lebih banyak pada kedua lokasi (khususnya jumlah lebih banyak terdapat pada lokasi Sendang Buleleng). Banyaknya cendawan ini ditemukan di lokasi Sendang Buleleng, karena cendawan ini lebih mampu berkompetisi di lingkungan tersebut. Hasil analisis tanah di Buleleng menunjukan hasil yang lebih baik dibandingkan di Karangasem, hal ini juga dapat mempengaruhi dominasi cendawan ini lebih banyak di lokasi Buleleng.
Menurut Syarief dkk., (2003) kompetisi yang dilakukan oleh cendawan ini yaitu dengan cara mengeluarkan metabolit sekunder yang bisa menghambat pertumbuhan cendawan lainnya, sehingga cendawan ini dapat mengabsorbsi nutrisi yang lebih banyak, dan menyebabkan pertumbuhannya lebih cepat. Menurut Waty (2012) ketersediaan unsur hara C, N, dan K yang rendah, tetapi Aspergilus niger mampu hidup dikarenakan cendawan ini mampu melarutkan posfat sebagai sumber nutrisi bagi cendawan ini dan tanaman. Menurut Pitt and Hocking (2006) cendawan yang termasuk dalam genus Aspergillus memerlukan temperatur yang lebih tinggi (450C), tetapi mampu
beradaptasi pada aw (activity water (0,8 aw) yang lebih rendah, sehingga mampu berkembang lebih cepat dibandingkan dengan Penicillium.
Berdasarkan hasil penelitian pada Tabel 2 cendawan Botrytis cinera ditemukan pada kedua lokasi yaitu di Karangasem yaitu dengan total (31 koloni), dan di Buleleng (11 koloni). Perbedaan jumlah cendawan ini yang ditemukan pada kedua lokasi dapat disebabkan karena perbedaan lokasi yang meliputi perbedaan lingkungan, ketersediaan unsur hara (C, N, P, K), dan dapat disebabkan pada saat sampling di lokasi Karangasem spora cendawan ini lebih banyak dari pada di Buleleng. Menurut Williamson dkk., (2007) secara umum cendawan Botrytis cinera dapat menyebabkan penyakit Grey mold pada tanaman. Bagian tanaman yang terkena cendawan ini akan menjadi nekrosis dan berubah warna dari kecokelatan hingga kehitaman.
Berdasarkan hasil penelitian pada Tabel 2 cendawan Cladosporium sphaeospermum ditemukan pada kedua lokasi yaitu pada lokasi Karangasem dengan total (6 koloni), sedangkan di Buleleng (8 koloni). Hal ini karena meratanya penyebaran spora cendawan ini di tanah pada kedua lokasi tersebut dan
cendawaan ini bersifat kosmopolit. Menurut Gandjar (1999) spesies ini memiliki sebaran luas dan berperan sebagai penyerangan sekunder pada semua tanaman. Spesies ini telah diisolasi dari tanah, udara, dan biji-bijian.
Cendawan Fusarium sporotrichioides hanya ditemukan pada lokasi di Sendang Buleleng dengan total (2 koloni), sedangkan di lokasi Sukadana Karangasem tidak ditemukan. Tidak ditemukannya cendawan ini di Sukadana Karangasem, dapat disebabkan karena sporanya tidak tersebar, kemungkinan cendawan ini tidak mampu berkompotisi pada lingkungan yang lebih ekstrim, serta ketersediaan unsur hara dan keadaan lingkungan di Sukadana Karangasem. Berdasarkan hasil analisis tanah menunjukan bahwa kadar air di Karangasem sangat rendah dibandingakn di Buleleng, sehingga pada lokasi ini tidak ditemukan cendawan ini. Berdasarkan hasil analisis tanah berkaitan dengan keberadaan cendawan Fusarium sporotrichioides pada lokasi Buleleng, karena kadar air di lokasi ini lebih banyak dibandingkan di Karangasem, sehingga cendawan ini mampu menyerap air melalui berkas pengangkut (xylem) dari tanaman mete yang dapat menunjang kehidupannya, diketahui bahwa cendawan
ini dapat menyebabkan penyakit layu pada tanaman.
Berdasarkan hasil penelitian pada Tabel 2 cendawan dari genus Penicilium ditemukan pada kedua lokasi yaitu di Sukadana Karangasem dan di Sendang Buleleng diantaranya Penicilium brevicompactum, P. citrinum, P. chrysogenum. Penicilium brevicompactum ditemukan pada kedua lokasi yaitu pada lokasi Karangasem (17 koloni) dan di Sendang Buleleng (32 koloni), hal ini karena penyebaran spora cendawan ini lebih banyak terdapat di Buleleng dibangdingkan di Karangasem. Cendawan ini bersifat kosmopolit yang tersebar luas di kedua lokasi tersebut. Menurut Scott et al., (2007) cendawan ini umumnya dapat diisolasi dari debu. Keberadan spesies ini pada kedua lokasi, karena dapat terkontaminasi dari debu maupun tanah, mampu mendegradasi serat selulose, dan material lain yang kaya akan selulose. Cendawan ini dapat memproduksi mikotoksin (Mycophenolic Acid (MPA) yang digunakan sebagai immunosuppressive.
Berdasarkan hasil penelitian pada Tabel 2 cendawan Penicilium citrinum hanya ditemukan pada lokasi Karangasem (29 koloni) sedangkan pada lokasi di Buleleng tidak ditemukan, hal ini karena
P. citrinum mempunyai kemampuan berkompetisi pada lingkungan yang lebih ekstrim (di Karangasem dibandingkan di Buleleng), selain itu penyebaran spora spesies ini sedikit, sehingga pada saat sampling kemungkinan sporanya tidak ikut terambil. Menurut Cayanto (2010) P.citrinum merupakan mirkoorganisme yang mempunyai pertumbuhan relatif cepat, serta mempunyai kemampuan menekan mikroorganisme lain (berkompetisi).
Berdasarkan penelitian pada Tabel 2 cendawan Penicilium chrysogenum ditemukan pada kedua lokasi yaitu di Sukadana Karangasem (5 koloni) dan di Sendang Buleleng (22 koloni), hal ini karena penyebaran spora cendawan ini lebih banyak terdapat di Buleleng dibangdingkan di Karangasem. Penyebaran spora cendawan ini meliputi keadaan dilingkungan pada lokasi Buleleng (berkaitan dengan hasil analisis tanah). Cendawan ini bersifat ini mampu hidup di tempat yang ekstrim dan bersifat kosmopolit (tersebar luas) di kedua lokasi tersebut. Spesies ini ditemukan pada kedua lokasi, karena mudah disiolasi dari rizosfer tanah, serasah dedaunan, serta di lingkungan (di dalam ruangan maupun di luar ruangan). Menurut Gandjar dkk., (1999) genus Penicilium dapat
menyebabkan penyakit pada tanaman dan diketahui sebagai penghasil metabolit sekunder telah digunakan dalam beberapa produk komersial kimia, termasuk antibiotik.
Berdasarkan hasil penelitian pada Tabel 2 cendawan Rhizopus stolonifer ditemukan pada kedua lokasi Karangasem dengan total (15 koloni) dan Buleleng (42 koloni). Pada lokasi Buleleng total koloni cendawan ini sangat banyak ditemukan, kemungkinan penyebaran spora ini sangat banyak dibandingkan pada lokasi Karangasem. Menurut Gandjar dkk., (1999) spesies ini tersebar luas di daerah tropis dan sub tropis, cendawan ini terdapat dalam tanah yang mengandung bahan organik, sehingga dapat mengalami proses pelapukan. Menurut Alexander (1976) Cendawan dari genus Rhizopus tergolong dalam kelompok hemiselulotik dan selulotik, karena Rhizopus dapat merombak lignin, lemak, selulosa, dan karbohidrat yang terdapat pada bahan organik tersebut.
Berdasarkan hasil penelitian pada Tabel 2 cendawan Synchephalastrum racemosum ditemukan pada kedua lokasi di Sukadana Karangasem (8 koloni) dan di Sendang Buleleng (6 koloni). Pada kedua lokasi ditemukannya spesies ini karena dapat berkompetisi pada lingkungan yang
berbeda dan dapat disiolasi dari tanah. Menurut Gandjar dkk., (1999) spesies ini bersifat saprofit, banyak ditemukan di daerah tropis maupun subtropis, yang mudah diisolasi dari tanah, dan serasah dedaunan. Spesies ini memiliki suhu pertumbuhan 170 - 400C.
Menurut Hyakumachi and Kubota (2003) cendawan tanah yang berada di rizosfer merupakan salah satu kelompok mikroba yang telah dilaporkan dapat menginduksi ketahanan tanaman terhadap berbagai penyakit, baik penyakit terbawa tanah maupun penyakit terbawa udara. Menurut Chanway (1997) keberadaan cendawan di rizosfer tanah dapat membantu pertumbuhan tanaman melalui berbagai mekanisme seperti: peningkatan penyerapan nutrisi, sebagai kontrol biologi terhadap serangan patogen, dan juga menghasilkan hormon pertumbuhan bagi tanaman. Menurut Baker and Cook (1974) secara alami tanah memiliki potensi mikroorganisme yang mampu menekan perkembangan patogen dalam tanah. Sebagian besar mikroorganisme antagonis tersebut hidup sebagai saprofit. Kemampuan organisme dalam beradaptasi terhadap berbagai keadaan lingkungan merupakan potensi besar untuk digunakan sebagai agen pengendali hayati. Menurut Waksman, (1952) keberadaan
mikroorganisme antagonis pada daerah rizosfer dapat menghambat persebaran dan infeksi akar oleh patogen, keadaan ini disebut hambatan alamiah mikroba. Mikroba antagonis sangat potensial dikembangkan sebagai agen pengendalian hayati. Selain sebagai agen antagonis, mikroorganisme tanah juga dapat mempengaruhi pertumbuhan tanaman dengan memproduksi senyawa-senyawa stimulat pertumbuhan seperti auksin dan fitohormon.
Hasil perhitungan pola penyebaran pada keanekaragaman jenis cendawan di Sukadana Karangasem mendapatkan hasil (0,85), angka tersebut berarti penyebarannya seragam. Menurut (Michael (1994) penyebaran teratur atau seragam adalah jumlah spesies yang terdapat pada tempat tertentu dalan komunitas. Penyebaran seragam terjadi apabila ada persaingan yang keras, sehingga menimbulkan kompotisi yang mendorong pembagian ruang hidup yang sama. Hasil perhitungan pola penyebaran cendawan di lokasi Sendang Buleleng mendapatkan hasil (1,03). angka tersebut berarti penyebarannya mengelompok. Menurut Michael (1994) penyebaran secara mengelompok merupakan pola yang sering ditemukan di alam, karena beberapa spesies tertentu yang hidupnya
selalu ada dalam kelompok-kelompok untuk mencakup kebutuhannya yang disebabkan oleh faktor lingkungan tekstur tanah, ketersediaan unsur hara, pH, kelembaban, dan ketersediaan air.
Hasil penelitian pada penyebaran cendawan di tanah baik secara seragam maupun mengelompok, hal tersebut dapat disebabkan karena pengaruh dari lingkungan pada kedua lokasi pengambilan sampel. Rao (1994) semua faktor lingkungan yang mempengaruhi penyebaran mikroorganisme (cendawan) dalam tanah. Banyaknya jumlah cendawan di dalam tanah dipengaruhi oleh banyaknya kandungan bahan organik, oksigen dan karbondioksida dalam atmosfer tanah pada kedalaman yang berbeda-beda.
Menurut Isroi (2006) perbedaan pada kedua lokasi, terutama dipengaruhi
oleh tekstur tanah. Perbedaan tekstur tanah pada kedua lokasi tersebut meliputi ketersediaan unsur hara yang terkandung di dalamnya, sehingga dapat mempengaruhi kesuburan tanah. Kesuburan tanah sangat berpengaruh terhadap keberadaan mikroorganisme dalam tanah (khususnya cendawan tanah) tergantung terhadap ketersediaan unsur-unsur yang dapat dimanfaatkan untuk menunjang kehidupannya. Kesuburan tanah sangat berpengaruh terhadap keberadaan mikroorganisme pada tanah, karena dapat berperan untuk menghancurkan limbah organik, fiksasi nitrogen, pelarutan fosfat, merangsang pertumbuhan, membantu penyerapan unsur hara, dan hasil perombakan dari mikroorganisme dalam tanah dapat dimanfaatkan oleh tanaman.
5
6
7
8
15
18
22
23
19
............—
|
No |
Keterangan Gambar |
No |
Keterangan Gambar |
|
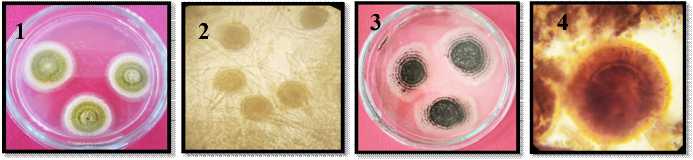
1. |
Makroskopis A. flavus |
13. |
Mikroskopis F. sporotrichoides |
|
2. |
Mikroskopis A. flavus |
14. |
Mikroskopis F. sporotrichoides |
|
3. |
Makroskopis A. niger |
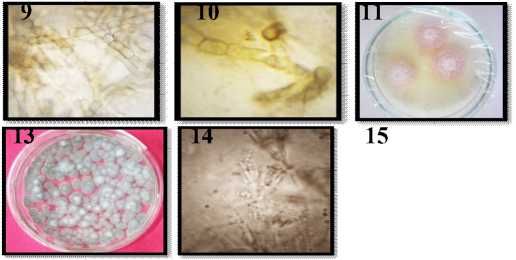
15. |
Makroskopis P. brevicompactum |
|
4. |
Mikroskopis A. niger |
16. |
Mikroskopis P. brevicompactum |
|
5. |
Makroskopis A. parasiticus |
17. |
Makroskopis P. citrinum |
|
6. |
Makroskopis A. parasiticus |
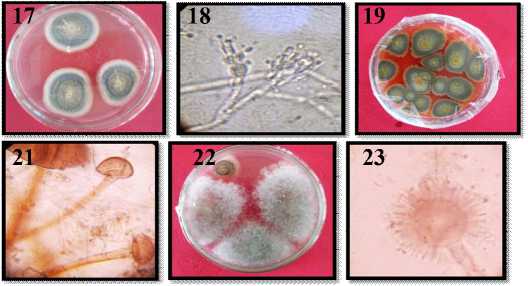
18. |
Mikroskopis P. citrinum |
|
7. |
Makroskopis Botrytis cinera |
19. |
Makroskopis P. chrysogenum |
|
8. |
Mikroskopis B. cinera |
20. |
Mikroskopis P. chrysogenum |
|
9. |
Makroskopis C. sphaeospermum |
21. |
Makroskopis R. stolonifer |
|
10. |
Mikroskopis C. sphaeospermum |
22. |
Mikroskopis R. stolonifer |
|
11 |
Mikroskopis C. sphaeospermum |
23. |
Makroskopis S. racemosum |
|
12. |
Makroskopis F. sporotrichoides |
24. |
Mikroskopis S. racemosum |
1. Rata-rata total koloni cendawan di Sukadana Karangasem adalah 32,6 x 103 CFU/g - 35 x 103 CFU/g, sedangkan di Sendang
Bulelelng 22,6 x 103 CFU/g -37,6 x 10-3 CFU/g.
-
2. Cendawan yang teridentifikasi pada rizosfer mete di kedua lokasi (Karangasem dan Buleleng) yaitu sebanyak 11 spesies. Lokasi Sukadana Karangasem terdapat sembilan 9 spesies yaitu Aspergilus niger, A. flavus, Botrytis cinera, Cladosporium sphaeospermum, Penicilium brevicompactum, P. citrinum, P. chrysogenum, Rhizopus stolonifer, dan Synchephalastrum racemosum. Lokasi Sendang Buleleng juga terdapat Sembilan 9 spesies yaitu Aspergilus niger, A. parasiticus, Botrytis cinera, Cladosporium sphaeospermum, Fusarium sporotrichoides, Penicilium brevicompactum, P.
chrysogenum, Rhizopus
stolonifer, dan Synchephalastrum racemosum.
-
3. Tingkat penyebaran jenis-jenis cendawan di Sukadana Karangasem adalah tergolong seragam (0,85) dan di Sendang Buleleng tergolong
mengelompok (1,03).
Saran
-
1. Diperlukan penelitian lebih lanjut untuk eksplorasi jenis-jenis cendawan tanah secara periodik, sehingga dapat diketahui jenis cendawan apa yang lebih dominan di daerah tersebut.
Daftar Pustaka
Abbas, H. K. 2005. Aflatoxin and Food Safety. CRC Press, Taylor & Francis Group: London.
Agrios, G.N. 1996. Ilmu Penyakit Tumbuhan. Edisi 3 (tiga). Terjemahan oleh Munsir Busnia, Gadjah Mada Umniversity Press. Yogyakarta.
Altas Indonesia dan Dunia. 2007. Edisi 33. Pustaka Agung Harapan. Surabaya.
Baker KF, RJ Cook. 1974. Biological Control of Plant Pathogens. WH. Freeman: San Francisco.
Barker, K.R. and W.W. Weeks. 1991. Relationships between soil and levels of Meloidogyne incognita and tobacco yield and quality. Journal of Nematology 23(1): 8290
Cayanto, D. 2010. Uji Mikroba
Aspergillus niger dan Penicillium
citrinum Sebagai Mikroba
Antagonis terhadap Patogen
Embun Tepung (Podosphaera leucotricha) Tanaman Apel Secara in vitro. http://www.shvoong.com.
Chanway, C.P. (1997). Inoculation of Tree Roots with Plant Growth Promoting
Bacteria: An Emerging technology for reforestation,Forest Science.
Darmawijaya,M. 1990. Klasifikasi Tanah. Gadjah Mada University Press. Yogyakarta.
Gandjar, I., R. A. Samson, K. T. Vermeulen, A. Oetari. I. Santoso. 1999. Pengenalan Kapang Topik Umum. Yayasan Obor Indonesia. Jakarta.
Gandjar, Indrawati & Wellyzar Sjamsuridzal. 2006. Mikologi Dasar dan Terapan. Yayasan Obor Indonesia, Jakarta.
Hanafiah. K. A., Iswandi A., A. Napoleon dan Nuni G. 2005. Biologi Tanah dan Limbah. Jakarta.
Hedayati, M. T., Pasqualotto, A. C., Warn, P. A., Bowyer, P., Denning. D. W. 2007. Aspergillus flavus : human pathogen, allergen, and mycotoxin producer. Microbiology 153: 16771692.
Hyakumachi, M and M Kubota. 2003. Fungi as plant growth promoter and disease suppressor. Pp. 101110 In: Fungal Biotechnology in Agricultural, Food and Environmental Application. Arora D. K. (ed) Marcel Dekker.
Isroi. S. 2006. Penelitian Mikroba. Balai Pertanian Bioteknology. Bogor. http://Ipardboo@Indo.net.id;Isroi@ Ipard.co [ 10 April 2007]
Jeger MJ. 2001. Biotic interaction and plant-pathogen association. In: Jeger MJ, Spence NJ. Biotic Interaction in Plant. Pathogen Association. CABL publishing: New York (USA).
Kawuri, R., Y. Ramona., I.B.G Darmayasa. 2007. Penuntun Praktikum Mikrobiologi Umum Untuk Study Farmasi F MIPA UNUD. Laboratorium
Mikrobiologi Jurusan Biologi F MIPA Universitas Udayana. Bukit-Jimbaran
Laboratorium Ilmu Tanah. 2011. Fakultas Pertanian. UNUD. Denpasar-Bali.
Michael, P. E., 1994. Metode Ekologi untuk Penyelidikan Ladang dan
Laboratorium.Universitas Indonesia, Jakarta.
Novriani dan A. Madjid. 2009. Dasar-dasar Ilmu Tanah. Prospek Pupuk Hayati Mikoriza. Bahan Kuliah Untuk Mahasiswa Fakultas Pertanian. Universitas Brawijaya Perss: Malang.
Odum, E.P, 1993. Dasar-dasar ekologi. Edisi ketiga. Gadjah Mada University Press.Yogyakarta.
Pitt, J. I., A. D. Hocking. 2006. Penicillium and related genera. Di dalam: C. W. Blackburn (ed). Food Spoilage Microorganisms.
Woodhead: CRC Press.
Proborini, M. W. 2002. Penuntun Praktikum Mikologi. Laboratorium Taksonomi Tumbuhan dan Mikologi Jurusan Biologi Fakultas Matematika Dan Universitas Udayana . Bukit Jimbaran.
Purwaningsih, Sri. 2005. Isolasi, Enumerasi, dan Karakterisasi Bakteri Rhizobium dari Tanah Kebun Biologi Wamena, Papua. Jurnal Biodiversitas.Vol.6(2)82-
84.
Rao, N.S.S. 1994. Mikroorganisme Tanah dan Pertumbuhan Tanaman Edisi kedua Penerbit Universitas Indonesia, UI Press. Jakarta.
Rukmana, Rachmad dan Saputra. 1997. Penyakitpenyakit tanaman
Hortikultura dan Teknik Pengendalian. Kanisius
:Yogyakarta.
Scheidegger ,K. A., Payne, G. A. 2003. Unlocking the secrets behind secondary metabolism: a review of Aspergillus flavus from pathogenicity to functional genomics. Journal Toxicol. 22: 423 – 459.
Scott, J.A. Bess Wong, Richard C. Summerbell, and Wendy A. Untereiner. 2007. A. survey of Penicillium brevicompactum and
P. biolowiezense from indoor environments, with commentary on the taxonomy of the P.brevicompactum group1. Botany 86: 732-741
Syarief, R., L. Egad an C.C Nurwitri. 2003. Mikotoksin Bahan Pangan. Institut Pertanian Bogor Press. Bogor.
Tarigan, Jeneng. 1988. Pengantar Mikrobiologi. : Depdiknas. Jakarta.
Wahyuaskari 2010. Habitat Mikroba Di Tanah
Available :
http://wahyuaskari.wordpress.com/ akademik/tanah-sebagai-habitat mikroorganisme/pengaruh-mikroba-terhadap-pertumbuhan-tanaman/
Opened : 01.06.2013
Waksman SA. 1952. Soil Mikrobiology. John Willey & John: New York.
Waty. R. 2012. Potensi Aspergillus niger dan Penicillium spp. Sebagai Endosimbion Pelarut Fosfat Pada Akar Serealia (Skirpsi). Institut Pertanian Bogor: Bogor.
Williamson B., B. Tudzynski, P. Tudzynski, J.A. Kan. 2007 Botrytis cinerea: The Cause of Grey Mould disease. Mold Plant Pathol. 8(5):561-580
Discussion and feedback