PENGAMATAN MORFOLOGI DAN ANATOMI BIBIT KAMBOJA JEPANG (Adenium sp.) AKIBAT PERENDAMAN BIJI DENGAN KOLKISIN
on
JURNAL SIMBIOSIS III (1): 322- 325
Jurusan Biologi FMIPA Universitas Udayana
ISSN: 2337-7224
Semtember 2015
PENGAMATAN MORFOLOGI DAN ANATOMI BIBIT KAMBOJA JEPANG (Adenium sp.) AKIBAT PERENDAMAN BIJI DENGAN KOLKISIN
MORPHOLOGICAL AND ANATOMICAL OBSERVATION OF DESSERT ROSE (Adenium sp.) SEEDLING AFTER SEED IMMERSION IN COLCHICINE
Putu Yunita Putri Aryani*dan Made Pharmawati
Jurusan Biologi FMIPA Universitas Udayana, Kampus Bukit Jimbaran, Bali
E-mail: yunitaputriaryani@gmail.com
INTISARI
Penelitian ini bertujuan untuk mengetahui pengaruh pemberian kolkisin melalui perendaman biji pada bibit kamboja jepang (Adenium sp.) dengan mengamati karakter morfologi dan anatominya. Penelitian ini dilakukan dengan perlakuan konsentrasi kolkisin 0%, 0,05%, 0,1%, dan 0,15%. Setiap perlakuan diulang 10 kali. Parameter yang diamati meliputi munculnya bibit, tinggi bibit, jumlah daun, panjang daun, lebar daun, dan kerapatan stomata. Hasil penelitian menunjukkan bahwa terjadi penghambatan munculnya bibit ke atas tanah akibat perlakuan perendaman dengan kolkisin. Tinggi bibit hasil perlakuan kolkisin lebih rendah dibandingkan dengan tinggi bibit kontrol. Secara anatomi pemberian kolkisin menyebabkan berkurangnya jumlah stomata pada kotiledon.
Kata kunci: anatomi, kamboja jepang, kolkisin, morfologi
ABSTRACT
This research aimed to determine the effect of colchicineby seed immersion ondessert rose (Adenium sp.) seedling. Observation were done on morphological and anatomical characters. This research was conducted using colchicine concentration of 0%, 0.05%, 0.1%, and 0.15%. Each treatment had 10 replications. The parameters observed included seedling emergence, seedling height, number of leaves, leaf length, leaf width, and stomatal density of cotiledone. The results showed differences in morphological characters, led to the emergence of seeds on the ground inhibited by immersion in colchicine. Anatomically giving of colchicines cause a reduction instomatal density of cotiledone.
Keywords: anatomy, colchicine, dessert rose, morphology
PENDAHULUAN
Kamboja jepang merupakan salah satu jenis tanaman hias yang berasal dari gurun Afrika dan Arab. Melihat dari asalnya, tanaman ini memerlukan sinar matahari penuh dan jenis media tanam yang porous. Tanaman kamboja jepang termasuk dalam tanaman semak sukulen yang tahan terhadap lingkungan yang kering dan panas (Sugih, 2005). Di Indonesia tanaman hias ini sangat cocok untuk dikembangkan karena Indonesia memiliki iklim tropis yang cenderung panas (Megawati, 2011).
Variasi warna bunga tanaman Adenium sp. banyak digemari oleh masyarakat Indonesia. Keindahan tanaman ini tidak hanya dari bunganya, tetapi juga dari akar dan batangnya yang berbentuk bonggol. Bentuk bonggolnya yang unik dan menarik mampu memikat para pecinta tanaman hias (Beikram dan Andoko, 2004). Haryanto (2005) menyatakan bahwa keindahan bonggol tanaman Adenium sp. adalah salah satu nilai jual dari tanaman ini. Bonggol merupakan kemampuan pangkal batang dan akarnya yang membesar. Umur mempengaruhi ukuran bonggol, semakin tua umur tanaman ini semakin besar ukuran bonggolnya.
Tanaman Adenium sp. memiliki variasi yang beragam. Keragaman genetis tanaman dapat ditingkatkan dengan hibridisasi (persilangan), seleksi mutan, introduksi, pembentukan poliploid, dan transfer gen (Poespodarsono, 1988; Lakitan, 1995). Tanaman poliploid merupakan tanaman yang memiliki lebih dari dua set kromosom dalam sel-selnya. Bagian tanaman poliploid lebih kekar dan besar meliputi akar, batang, daun, bunga, dan buah dibandingkan dengan tanaman diploid. Hal ini merupakan sifat umum dari tanaman poliploid (Suryo, 1995). Induksi poliploid sering dimanfaatkan dalam pemuliaan tanaman karena hasil panen menjadi lebih baik (Alam et al., 2011).
Poliploid dapat terjadi secara alami maupun buatan pada tanaman. Poliploid secara buatan dapat dilakukan dengan zat kimia, salah satunya dengan kolkisin. Kolkisin adalah zat kimia yang diekstrak dari umbi dan biji tanaman Autumn crocus (Colchicum autumnale). Zat ini
menyebabkan poliploid dengan menghambat pembentukan benang-benang spindel (mikrotubul) saat pembelahan sel sehingga terjadi penggandaan kromosom (Hess, 1975; Poespodarsono, 1988). Setiap jenis tanaman memiliki perbedaan respon terhadap pemberian kolkisin (Suryo, 1995). Umumnya kolkisin akan bekerja secara efektif pada konsentrasi 0,01 – 1% dan lama perlakuan antara 6-72 jam (Suminah dan Setyawan, 2005). Pengaruh berbagai konsentrasi kolkisin terhadap tanaman perlu diketahui, sehingga diperoleh konsentrasi optimum untuk mendapatkan tanaman poliploid. Penelitian ini bertujuan menguji pengaruh konsentrasi kolkisin pada tanaman Adenium sp.
MATERI DAN METODE
Bahan Tanaman
Biji tanaman kamboja jepang diperoleh di kebun tanaman hias di Denpasar Bali. Biji kamboja jepang (Adenium sp.) yang digunakan berasal dari buah yang sudah tua. Selanjutnya dilakukan uji viabilitas biji dengan merendam biji dalam air dan dipilih biji yang tenggelam. Biji yang terseleksi selanjutnya dikecambahkan pada petridish.
Perlakuan dengan Kolkisin
Konsentrasi kolkisin yang digunakan adalah 0,05%, 0,1%, dan 0,15%. Konsentrasi tersebut dibuat dengan melakukan pengenceran dari stok 0,2 %. Sebagai kontrol digunakan akuades. Kecambah dengan panjang 0,5 cm selanjutnya direndam dalam kolkisin selama enam jam masing-masing sepuluh kecambah tiap perlakuan termasuk kontrol. Setelah perendaman kecambah dibilas dengan air untuk menghilangkan kolkisin yang masih menempel.
Kecambah ditanam dalam polybag dalam medium tanah. Penyiraman dilakukan sekali sehari. Pengamatan dilakukan selama 33 hari setelah tanam (HST).
Pengamatan dan Analisis Data
Pengamatan morfologi tanaman kamboja jepang meliputi tinggi bibit, panjang dan lebar kotiledon, serta panjang dan lebar daun. Pengamatan morfologi ini
dilakukan dengan cara manual dengan menggunakan penggaris dengan satuan cm.
Karakter anatomi yang diamati pada bibit kamboja jepang yaitu jumlah stomata pada kotiledon tanaman. Kotiledon tanaman direndam dengan menggunakan pemutih pakaian konsentrasi 10% selama 30 menit sehingga berubah warna menjadi putih. Stomata kemudian diamati di bawah mikroskop pada perbesaran objektif 20x dan okuler 10x atau perbesaran 200x. Dihitung jumlah stomata pada bidang pandang. Data yang diperoleh dianalisis dengan ANOVA menggunakan Rancangan Acak Kelompok.
0.9 0.8
'E 0.7
~ 0.6
'S 0.5 ⅞ 0.4 CiD 0.3 ⅛ 0.2
CL 0.1 0
-0.1
a
→-0%
-■-0.05%
-⅛-0.10%
—•-0.15%
7 HST 21 HST 35 HST
Waktu
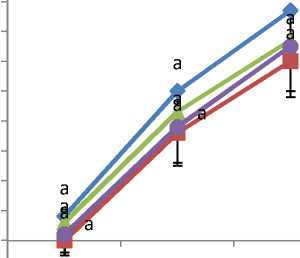
Gambar 2. Rata-rata Panjang Kotiledon Akibat
Pemberian Kolkisin
HASIL
Pengamatan Morfologi Bibit Kamboja Jepang (Adenium sp.)
Pengaruh pemberian kolkisin dengan konsentrasi 0,05%, 0,1%, dan 0,15% terhadap munculnya bibit ke atas tanah dalam satuan persen (%) dapat dilihat pada Tabel 1.
Tabel 1. Persentase Munculnya Bibit ke Atas Tanah (%) Persentase Munculnya Bibit Ke Atas Tanah
|
Waktu Konsentrasi |
6 HST |
8 HST |
|
0% |
100,0 ± 0,00a |
100 ± 0a |
|
0,05% |
53,00 ± 14,0bc |
100 ± 0a |
|
0,10% |
80,67 ± 9,93ab |
100 ± 0a |
|
0,15% |
38,67 ± 5,67c |
100 ± 0a |
→-0%
-■-0.05%
-⅛-0.10%
—•-0.15%
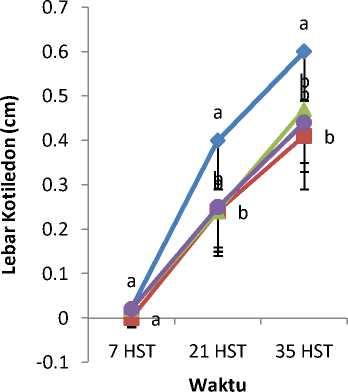
Gambar 3. Rata-rata Lebar Kotiledon Akibat
Pemberian Kolkisin
Bibit tanaman adenium pada 6 HST dengan konsentrasi kolkisin 0% tidak berbeda nyata pada bibit tanaman adenium 8 HST. Perlakuan dengan konsentrasi kolkisin 0,05%, 0,10%, dan 0,15% 6 HST memberikan pengaruh yang berbeda nyata pada tanaman adenium 8 HST.
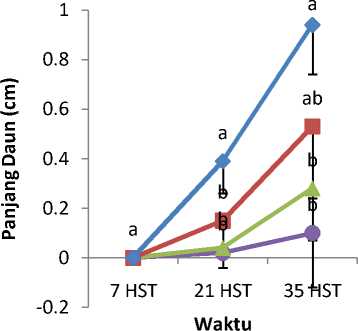
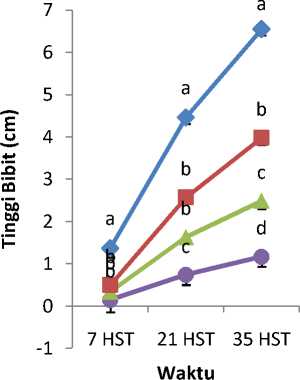
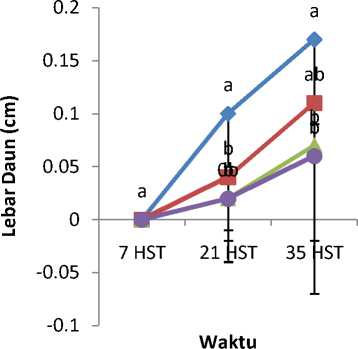
Mutagen kolkisin dengan konsentrasi berbeda mempengaruhi tinggi tanaman (Gambar 1), panjang kotiledon (Gambar 2), lebar kotiledon (Gambar 3), panjang daun (Gambar 4), dan lebar daun (Gambar 5). Tanaman
→-0%
-■-0.05%
-⅛-0.10%
—•-0.15%
kontrol lebih tinggi dibandingkan dengan tanaman yang diberi perlakuan kolkisin.Panjang kotiledon dan lebar kotiledon tertinggi yaitu pada bibit tanaman kontrol. Tanaman kontrol juga memiliki panjang daun dan lebar daun yang paling tinggi.
→-0%
-■-0.05%
-⅛-0.10%
-•-0.15%
Gambar 1. Rata-rata Tinggi Bibit Akibat Pemberian
Kolkisin
Gambar 4. Rata-rata Panjang Daun Akibat Pemberian Kolkisin
→-0%
-■-0.05%
-⅛-0.10%
—•-0.15%
Gambar 5. Rata-rata Lebar Daun Akibat Pemberian
Kolkisin
Jumlah daun paling banyak pada tanaman kontrol. Semakin tinggi konsentrasi kolkisin yang diberikan, semakin sedikit jumlah daun yang dihasilkan oleh tanaman tersebut. Hal ini ditunjukkan pada Tabel 2. Pada perlakuan kolkisin konsentrasi 0,05%, 0,10%, dan 0, 15% memiliki rata-rata jumlah daun yang berbeda nyata dengan kontrol.
Tabel 2. Rata-rata Jumlah Daun Usia 33 HST
Rata-rata Jumlah Daun 33 HST
|
Konsentrasi |
Rata-rata Jumlah Daun |
|
0% |
1,9 ± 0,23a |
|
0,05% |
1 ± 0,33b |
|
0,10% |
0,7 ± 0,34bc |
|
0,15% |
0,3 ± 0,22c |
Pengamatan Anatomi Bibit Kamboja Jepang (Adenium sp.)
Pengamatan anatomi tanaman kamboja jepang dengan mengamati jumlah stomata pada masing-masing perlakuan. Tabel 3 menunjukkan rata-rata jumlah stomata pada kotiledon bibit kamboja jepang pada masing-masing ulangan. Data yang diperoleh adalah rata-rata jumlah stomata pada bibit dengan konsentrasi kolkisin 0,05%, 0,10%, dan 0, 15% tidak berbeda nyata dengan jumlah stomata pada bibit kontrol.
Tabel 3. Rata-rata Jumlah Stomata Bibit Adenium per Ulangan.
Rata-rata Jumlah Stomata per Ulangan
|
Konsentrasi |
Rata-rata Jumlah Stomata |
|
0% |
9 ± 6,08a |
|
0,05% |
7,67 ± 2,31a |
|
0,10% |
8 ± 4,36a |
|
0,15% |
2 ± 1,73a |
PEMBAHASAN
Perlakuan dengan konsentrasi kolkisin 0,05%, 0,10%, dan 0,15% 6 HST memberikan pengaruh yang berbeda nyata pada munculnya bibit tanaman adenium 8 HST. Bibit tanaman kontrol lebih cepat muncul, dibandingkan bibit yang berasal dari biji yang direndam dengan kolkisin. Pada 6 HST, persentase munculnya bibit akibat pengaruh kolkisin tidak memperlihatkan trend yang berhubungan dengan konsentrasi kolkisin. Hal itu dapat disebabkan karena kolkisin tidak mempengaruhi keseluruhan sel atau mengakibatkan mutasi sel secara acak. Sel-sel yang sedang aktif membelah akan peka terhadap kolkisin, sedangkan sel-sel yang telah terdiferensiasi kurang peka terhadap mutagen ini.
Perbedaan fisiologis masing-masing biji juga berpengaruh terhadap kepekaan terhadap kolkisin. Hal ini sesuai dengan Byrd (1983) yang meyatakan bahwa tingkat kemasakan biji berhubungan erat dengan kemampuan biji untuk tumbuh. Biji yang belum masak memiliki kemampuan tumbuh dan berkembang yang kurang baik dibandingkan dengan biji yang telah masak meskipun biji yang belum masak tersebut mampu mengalami perkecambahan.
Pada 8 HST, semua bibit dengan perlakuan kolkisin mucul ke atas tanah karena kolkisin merupakan mutagen yang bersifat racun bagi tanaman. Mutagen ini bekerja pada jaringan meristematik dari tanaman, dimana kolkisin menghambat aktifnya enzim-enzim dan hormon untuk pertanaman. Hal tersebut terjadi karena adanya aberasi kromosom yang disebabkan oleh kolkisin (Sheeler dan Bianchi, 1987). Mekanisme asimilasi terganggu akibat pemberian kolkisin pada tanaman, dengan demikian pertanaman kecambah dengan perlakuan kolkisin perlu waktu lama karena kecambah memerlukan waktu untuk beradaptasi (Roychowdhury dan Tah, 2011).
Pengamatan morfologi bibit adenium selama 33 HST menunjukkan bahwa tanaman kontrol mengalami
pertanaman lebih cepat dalam beberapa parameter yang diamati. Kolkisin mempengaruhi pertanaman tanaman adenium dengan mempengaruhi penyusunan mikrotubula dalam sel. Setiap dimmer penyusun mikrotubula (gabungan a dan b tubulin) mempunyai tempat pengikat spesifik bagi kolkisin. Dimmer yang telah berikatan dengan kolkisin akan mencegah dimmer tersebut tersusun menjadi mikrotubula. Apabila dimmer yang terhalangi penyusunannya adalah mikrotubula yaitu penyusun benang spindel pada mitosis, akan menyebabkan pembelahan sel terhambat dan kromosom yang telah mengganda selama interfase gagal memisahkan diri, sehingga membentuk sel yang poliploid (Sheeler dan Bianchi, 1987).
Selain menyusun benang-benang spindel, mikrotubula bersama mikrofilamen dan filamen intermediet juga menyusun rangka sel yang berperan dalam proses transportasi intraseluler molekul kecil seperti air dan ion anorganik pada sitoplasma (Kimball, 1995). Kolkisin dapat menghalangi perakitan mikrotubula penyusun rangka sel dan mengacaukan tata letak beberapa protein pada membran sel khususnya glikoprotein yang diatur oleh mikrotubula dan mikrofilamen yang berperan sebagai protein reseptor dan pengikat milekul lain pada permukaan sel (Wolfe, 1983). Rusaknya formasi mikrotubula dalam sel menyebabkan gangguan transportasi sel, menyebabkan molekul-molekul dalam sitoplasma tidak terdistribusi dengan baik. Pertanaman tanaman menjadi lambat setelah benih diberikan perlakuan kolkisin pada konsentrasi tinggi.
Parameter jumlah daun menunjukkan bahwa kolkisin menyebabkan jumlah daun lebih sedikit dibandingkan kontrol. Pembelahan sel yang lambat juga menyebabkan pertanaman dan perkembangan primordial daun menjadi lambat. Daun merupakan organ fotosintesis yang utama sehingga menentukan jumlah asimilat yang dihasilkan selama proses pertanaman dan perkembangan (Kimball, 1995).
Pengamatan anatomi bibit adenium dilakukan dengan pengamatan jumlah stomata pada kotiledon bibit masing-masing perlakuan kolkisin. Rata-rata jumlah stomata yang diperoleh pada bibit kontrol berjumlah paling banyak dibandingkan dengan tanaman yang diberikan perlakuan kolkisin. Bibit adenium dengan perlakuan konsentrasi kolkisin 0,15% memiliki rata-rata jumlah stomata terendah. Induksi poliploid oleh kolkisin dengan konsentrasi tinggi menyebabkan ukuran stomata menjadi lebih besar dan jumlah stomata pada satu bidang pandang perbesaran 40x menjadi menurun. Tanaman yang bersifat poliploid secara anatomi dapat ditandai dengan ukuran sel-selnya menjadi lebih besar, terlihat jelas pada sel epidermis, inti sel, dan stomata (Suryo, 1995).
Pada tanaman poliploid, jumlah sel penjaga lebih sedikit tetapi memiliki ukuran sel penjaga lebih besar dibandingkan tanaman diploid dengan kandungan kloroplas yang lebih banyak. Jumlah kloroplas pada sel penjaga dapat menunjukkan tingkat ploidi. Lozykowska (2003); Liu et al.,(2007); dan Omidbaigi et al., (2010) melaporkan bahwa ukuran stomata dapat mengindikasikan tingkat ploidi dimana semakin besar ukuran stomata semakin tinggi tingkat ploidi. Penelitian Ho et al. (1990); Lozykowska (2003) menyatakan bahwa ukuran panjang stomata berhubungan dengan jumlah kloroplas pada sel penjaga. Semakin panjang ukuran stomata maka semakin banyak jumlah kloroplas pada sel penjaga dan semakin tinggi tingkat ploidi.
SIMPULAN
Perlakuan kolkisin dengan konsentrasi 0,05%, 0,1%, dan 0,15% menyebabkan munculnya bibit ke atas tanah terhambat. Tinggi tanaman, panjang dan lebar kotiledon, jumlah daun tanaman perlakuan pada bibit hasil kolkisin lebih kecil dibandingkan bibit kontrol.Jumlah stomata pada kotiledon pada perlakuan kolkisin berjumlah lebih sedikit dibandingkan dengan bibit kontrol.
KEPUSTAKAAN
Alam, M.M., M.K. Karim, M.A. Aziz, M.M Hossain, B.
Ahmed, A. Mandal. 2011. Induction and evaluation of polyploidy in some local potato varieties of Bangladesh. J. BiodiversityEnviron. Sci. 1: 16-21.
Beikram dan A. Andoko. 2004. Mempercantik Penampilan Adenium. Agromedia Pustaka: Jakarta.
Byrd, K.W. 1983. Pedoman Teknologi Benih. Terj. Hamidin, E. PT.Pemimpin Masa: Jakarta.
Haryanto, E. T. 2005. Beberapa Teknik Peningkatan Nilai Estetika Tanaman Adenium.Seminar Nasional Agribisnis Tanaman Hias UNS. Surakarta: 1 Oktober 2006.
Hess, D. 1975. Plant Physiology: Biomolecular, Biochemical, and Physiologycal.Fundamental of Metabolism and development Springer-Velag New York Inc.New York. pp 144-159.
Ho, I., Y. Wan., J.M. Widhlom., and A.L. Rayburn. 1990.
The use of stomatal chloroplast number for rapid determination of ploidy level in maize. J.Pl.
Breed. 105(3): 203-210.
Kimball, J.W. 1995. Biologi. Jilid 2 edisi ke-5. Penerjemah Siti Soetarmi Tjitrosomo dan Nawangsari Sugiri. Penerbit Erlangga: Jakarta.
Lakitan, B,.1995. Pemuliaan Tanaman Secara In Vitro dengan Memanfaatkan Variasi Somaklonal: Prospek dan Masalahnya. Kumpulan Makalah
Seminar Bioteknologi dan Pelatihan Teknologi DNA. Fakultas Pertanian USU:Medan. Hal.1-9.
Liu, G., Z. Li., and M. Bao. 2007. Colchicine-induced chromosome doubling in Plantanus acefolia and its effect on plant morphology. Euphytica DOI 10.1007/s10681-007-9406-6.
Lozykowska, K.S. 2003. Determination of the ploidy level in chamomile (Chamomilla recutita (L.)Rausch.)strains rich in α-bisabol. J. Appl. Gent. 44(2): 151-155.
Megawati, A.S. 2011.Pengaruh Zat Pengatur Tumbuh Akar dan Media Tanam terhadapKeberhasilan dan Pertanaman Setek Kamboja Jepang (Adenium obesum). Skripsi. Fakultas Pertanian, Universitas Sumatera Utara: Medan.
Omidbaigi, R., M.E. Hassani., and S. Yavari. 2010. Induction od autotetraploidy in dragonhead (Dracocephalum moldavica L.) by colchicine treatment. J. Fruit & Ornmt. Pl. Res. 18(1): 23-35
Poespodarsono, S. 1988. Dasar-Dasar Ilmu Pemuliaan Tanaman. IPB: Bogor.
Roychowdhury R. and J. Tah.2011.Chemical mutagenic action on seed germination and related agro-metrical traits in M1 Dianthus generation. Curr. Bot. 2(8): 19-23.
Sheeler, P. and D.G. Bianchi., l987.Cell and Molecular Biology. John Wiley and Sons: Inc. Canada
Sugih, O. 2005.88 Variasi Adenium Agar Rajin Berbunga.
Penebar Swadaya: Jakarta.
Suminah S. dan A.D. Setyawan. 2005. Induksi poliploid bawang merah (Alliumascalonicum L.)dengan pemberian kolkisin. Biodeversitas 6(1): 174-180.
Suryo., l995. Sitogenetika. Gadjah Mada University Press: Yogyakarta.
Wolfe, S.L., l983. Introduction to Cell Biology.
Wadsworth Publishing Company:
California.
322
Discussion and feedback