ANALYSIS OF PATHOGENIC IUTA GENE MARKERS IN ESCHERICHIA COLI CAUSES OF COLLIBACILLOSIS IN FREE-RANGE CHICKEN
on
Volume 13 No. 1: 57-66
Pebruari 2021
DOI: 10.24843/bulvet.2021.v13.i01.p10
Buletin Veteriner Udayana
pISSN: 2085-2495; eISSN: 2477-2712
Online pada: http://ojs.unud.ac.id/index.php/buletinvet
Terakreditasi Nasional Peringkat 3, DJPRP Kementerian Ristekdikti No. 21/E/KPT/2018, Tanggal 9 Juli 2018
Analisis Marka Gen Patogenik iutA Escherichia Coli Penyebab Colibasilosis pada Ayam Buras
(ANALYSIS OF PATHOGENIC IUTA GENE MARKERS IN ESCHERICHIA COLI CAUSES OF COLLIBACILLOSIS IN FREE-RANGE CHICKEN)
Kadek Satria Adi Marhendra1*, I Gusti Ngurah Kade Mahardika2, I Nengah Kerta Besung3, I Gusti Ketut Suarjana3
1Mahasiswa Program Profesi Dokter Hewan, Fakultas Kedokteran Hewan, Universitas Udayana, Jl. PB. Sudirman Denpasar, Bali; 2Laboratorium Virologi Veteriner, Fakultas Kedokteran Hewan, Universitas Udayana, Jl. Raya Sesetan, Gg. Markisa No. 6, Denpasar, Bali; 3Laboratorium Mikrobiologi Veteriner Universitas Udayana, Fakultas Kedokteran, Universitas Udayana, Jl. PB. Sudirman Denpasar, Bali. *Email: kecik_kecil@yahoo.com
Abstrak
Kolibasilosis merupakan penyakit menular pada ayam yang disebabkan oleh Avian Pathogenic Escherichia coli (APEC). Kemampuan APEC untuk menyebabkan penyakit tergantung pada banyak faktor patogen, salah satunya adalah gen patogenik iutA. Penelitian bertujuan untuk mengetahui sekuen DNA gen iutA APEC di Bali serta kekerabatannya dengan gen iutA dari negara lain. Penelitian ini menggunakan dua isolat APEC asal ayam buras di Kabupaten Tabanan dan Badung yang telah dimurnikan dan tersedia di Laboratorium Bakteriologi Fakultas Kedokteran Hewan, Universitas Udayana. DNA isolat diisolasi dengan Chelex 10%. Produk polymerase chain reaction (PCR) disekuensing di First Base Laboratories Malaysia dengan metode Sanger’s dideoxy nucleotide termination. Kedua hasil sekuen gen iutA memiliki homologi 100% dengan panjang 250 bp. Analisis filogenik dengan 58 data gen iutA di Escherichia coli dan bakteri lain di dunia memiliki 43 situs polimorfik pada tingkat asam nukleat dan 13 di tingkat asam amino. Gen iutA asal Bali berada di dalam satu kelompok dengan gen iutA asal Brazilia (KP657535) tahun 2011, Jerman (LT599825) tahun 2016, dan Cina (CP033635) tahun 2016. Gen ini dapat digunakan sebagai marker patogenik APEC di Indonesia.
Kata kunci: Escherichia coli; APEC; gen; patogenik; iutA; filogenik
Abstract
Colibacillosis is an infectious disease in chickens caused by Avian Pathogenic Escherichia coli (APEC). The ability of APEC to cause disease depends on many pathogenic factors, one of which is the iutA pathogenic gene. This study was purposed to determine the DNA sequence of the APEC iutA gene in Bali and its kinship with the iutA gene from other countries. Two APEC isolates from free-range chicken in Tabanan dan Badung has been used. The isolates have been purified and were available at the Bacteriology Laboratory of Veterinary Medicine Faculty, Udayana University. Chelex 10% was used for DNA isolation. DNA amplification using published DNA primer has been conducted with the polymerase chain reaction (PCR) method. The PCR product was sequenced at Malaysia, First Base Laboratories using Sanger’s Dideoxy Nucleotide Termination method. The iutA gene of both isolates can be analyzed and have 250 bp in length. Both were 100% homologous. A phylogenic test using 58 DNA sequences of iutA gene from Escherichia coli and other bacteria in the world was conducted in MEGA 5.2. All data have 43 polymorphic sites of nucleotide acid and 13 polymorphic sites of amino acid. The iutA gene from Bali is in one group, with the iutA gene from Brazil (KP657535) in 2011, Germany (LT599825) in 2016, and China (CP033635) in 2016. This gene can be used as a pathogenic marker of APEC in Indonesia.
Keywords: Escherichia coli; APEC; gene; pathogenic; iutA; phylogenic.
PENDAHULUAN
Salah satu penyakit menular pada ayam adalah kolibasilosis yang disebabkan oleh bakteri Escherichia coli (E.coli) galur patogen atau Avian Pathogenic Escherichia coli (APEC). Infeksi penyakit ini dapat menimbulkan berbagai macam gangguan pertumbuhan dan dapat menyebabkan kematian (Tarmudji, 2003). Kolibasilosis menimbulkan kerugian pada peternakan ayam (Landman et al., 2014; Lau et al., 2012). Hal ini berdampak penting bagi peternakan ayam.
Kasus kolibasilosis telah dilaporkan di berbagai daerah di Indonesia. Penyakit ini telah ditemukan pada ayam petelur maupun pedaging di berbagai daerah (Tarmudji, 2003). Di Bali, penyakit ini ditemukan di peternakan ayam di berbagai kabupaten (Barus et al., 2013). Beberapa upaya telah dilakukan untuk mengatasi penyakit kolibasilosis salah satunya dengan pemberian antibiotik. Namun, pemberian antibiotik secara terus-menerus membuat kolibasilosis mudah mengalami resistensi terhadap antibiotik tertentu (Luhung et al., 2017).
Faktor gen virulensi APEC merupakan faktor yang menentukan kemampuan untuk menyebabkan penyakit. Beberapa gen ini ada di dalam plasmid. Dari beberapa gen yang ditemukan, lima gen yang umum yaitu iutA, hlyF, episomal iss, iroN, dan episomal ompT. Gen ini merupakan gen yang disebut sebagai marka patogenik dari APEC (Johnson et al., 2008). Tingkat frekuensi deteksi gen iutA berbeda-beda di berbagai negara. Gen iutA berasosiasi dengan sifat patogenis APEC, dimana frekuensi gen patogen iutA pada APEC di negara lain yaitu di Amerika Serikat sebesar 79,5% (Johnson et al., 2006), di Zimbabwe sebesar 80% (Mbangai dan Nyararai, 2015), di Cina sebesar 70,3% (Wang et al., 2015), dan di Eropa (Perancis, Spanyol, dan Belgia) sebesar 82,7% (Schouler et al., 2012). Gen iutA di E.coli galur patogen memiliki frekuensi gen iutA 79,5% dibandingkan dengan frekuensi gen iutA
pada E.coli komensal yang hanya 34,2% (Johnson et al., 2006). Frekuensi gen marka patogenik iutA APEC di Indonesia belum pernah dilaporkan.
Penelitian ini menganalisa gen patogenik iutA dari ayam buras asal Kabupaten Tabanan dan Badung, Provinsi Bali dan membandingkan gen iutA di Bali dengan gen iutA yang ada di seluruh dunia.
METODE PENELITIAN
Objek Penelitian
Sampel yang digunakan merupakan isolat E. coli yang berasal dari organ ayam buras penderita kolibasilosis dengan ciriciri bulu kusam, diare, nafsu makan menurun, kurus, pembengkakan pada daerah kepala, dan perdarahan pada jaringan subkutan. Isolat yang digunakan sebanyak dua isolat yang berasal dari Kabupaten Tabanan dan Kabupaten Badung. Isolat telah positif pada media Eoisn Metylen Blue Agar (EMBA), uji TSIA, SIM, Katalase, MR, Glukosa, Laktosa dan negatif dengan uji Oksidase, SCA dan VP. Isolat didapatkan dan dimurnikan di Laboratorium Bakteriologi, Fakultas Kedokteran Hewan, Universitas Udayana
Isolasi DNA
Isolasi DNA menggunakan metode Chelex (Walsh et al., 1991). Sampel E.coli dimasukan ke dalam tabung eppendorf yang sudah berisi larutan chelex 10%. Larutan itu vortex atau shaker selama 15 detik, kemudian dimasukan ke dalam sentrifuge selama 1 menit, dipanaskan selama 60 menit pada suhu 94oC. Prosedur diulang dua kali, lalu sampel disimpan selama satu malam pada suhu 4oC. Sampel siap digunakan untuk dilakukan tindakan PCR
Uji PCR
Gen diamflifikasi dengan
menggunakan sepasang DNA primer iutA : iutAF5’-
CGTCGGGAACGGGTAGAATCG -3’ untuk menghasilkan pita 302 bp (Johnson et al., 2006). DNA primer masing-masing sebanyak 0,5 µL. Metode uji PCR sesuai dengan yang dilakukan oleh Brown, 1992.
Program PCR dengan aturan suhu predenaturasi 94oC selama 7 menit, denaturasi 94oC selama 1 menit, suhu annealing 55oC selama 45 detik, dan ekstensi 75oC selama 1 menit 30 detik. Siklus tersebut dilakukan sebanyak 35 kali siklus. Setelah itu perpanjangan langkah terakhir (post ekstensi) 75oC selama 5 menit. Suhu annealing ditetapkan setelah ditentukan sebelumnya di lab Biomedik Fakultas Kedokteran Hewan, Universitas Udayana.
Sekuensing DNA
Hasil PCR APEC yang posistif iutA akan disekuensing DNA menggunakan metode Sanger dedeoksi termination melalui First Base Laboratories Malaysia.
Analisis Data
Hasil penelitian data sekuensing digunakan sebagai data primer. Data primer akan dilakukan analisis filogenik dan jarak genetik dengan data sekunder yang didapatkan dari gene bank dengan informasi asal negara dan kode akses (geneid). Analisis filogenik dan jarak genetik akan menggunakan program lunak analisis DNA yaitu MEGA 5.2 atau Molecular Evolutionary Genetics Analysis (Tamura et al., 2011).
HASIL DAN PEMBAHASAN
Hasil
Isolat E. coli asal Bali yang digunakan adalah isolat APEC dengan kode isolat E3 asal Tabanan dengan kode PCR A2984 dan isolat E7 asal Badung dengan kode PCR A2985 tahun 2018. Kedua sampel berhasil diamplifikasi dengan PCR. Hasil berupa visualisasi PCR gen iutA APEC asal ayam buras dengan panjang 300bp terlihat pada gambar 1. Hasil sekuensing kedua gen iutA yang dapat dianalisis dengan baik sepanjang 250 bp. Kedua sekuen tersebut menunjukkan urutan basa gen iutA yang
sama (homologi 100%). Hasil sekuensing gen iutA diregistrasi di genebank dengan kode akses (accession number) MK776781 dan MK776782.
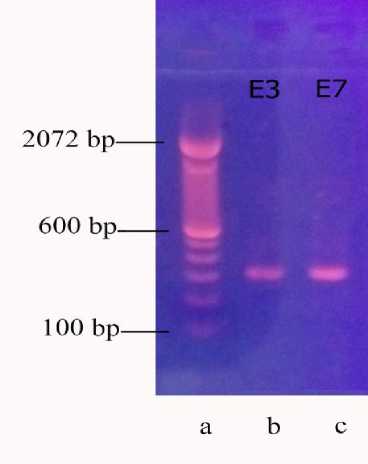
Gambar 1. Elektroforesis agarosa 1% (a) Ladder Invitrogen 100 bp, (b) sampel E3 (c) sampel E7, divisulasisasi pada UV dengan pewarna ethidium bromide.
Data primer dan 58 data sekunder yang diunduh dari genebank dianalisis menggunakan metode Uweighted Pair-Group Method with Arithmetic (UPGMA) tree dengan metode bootstrap (500 pengulangan). Hasil berupa pohon filogenik ditampilkan pada gambar 2. Gen iutA dari kode akses MK776781 dan kode akses MK776782 berada dalam satu kelompok yang sama dengan gen iutA asal Brazilia (KP657535) tahun 2011, Jerman (LT599825) tahun 2016, dan Cina (CP033635) tahun 2016. Secara filogeni gen iutA E.coli dunia dikelompokkan menjadi dua kelompok besar. Pembagian kelompok genetik tersebut mempunyai dukungan nilai bootstrap yang kuat (99%). Kelompok 1 sebelah atas selanjutnya terbagi menjadi dua yaitu 1A dan 1B, sedangkan yang sebelah bawah menjadi 2A dan 2B. Pembagian 1A dan 1B mempunyai dukungan bootstrap 87%, sementara 2A dan 2B 95%. Isolat yang diperoleh dalam penelitian ini termasuk klaster 1B.
87
LM996880 Norway 2010
LS992168 Germany 2018
MK111112 Iran 2012
CP027119 France 2010
KY293686 Russia 2016
CP019055 Hongkong 2014
CP028304 Pakistan 2016
CP031295 Vietnam 2017
CP019018 Argentina 2010
CP023826 Sweden 2009
CP035377 Brazil 2015
CP026854 Slovakia 2018
CP021179 United Emirates Arab 2009
CP009579 Netherlands 2014
CP023363 UK Scotland 2002
AP018784 Japan 2007
CP035124 China 2017
CP024827 South Korea 2015
99
M CP018994 Israel 2012
MG649062 Italy 2017
CP010372 Colombia 2012
MH422552 Spain 2015
CP023145 Egypt 2016
CP006784 Denmark 2013
MH753063 Malaysia 2018
CP035516 USA 2015
CP029978 Switzerland 2018
LR130556 Australia 2018
CP022069 S.enterica USA 2011
KP119165 Canada 2014
LC056228 India 2015
CP030791 USA 1982
CP022731 Saudi Arabia 2012
CP027851 Mexico 2010
CP026643 South Korea 2017
CP025403 Qatar 2016
CP031653 UK London 2016
rl MK776782 INDONESIA (BALI) 2018
■ MK776781 INDONESIA (BALI) 2018
M CP033635 China 2016
87
KP657535 Brazil 2011
LT599825 Germany 2016
CP011331 Denmark 2011
99
CU928145 France 2008
95 CP024992 Germany 2011
CP016828 South Korea 2008
CP025707 USA 2018
CP034067 Shigella sonnei USA 2015
CP021175 Taiwan 2016
CP012693 South Korea 2003
I CU928163 France 2008
CP014092 USA 2018
CP025573 Sweden 2008
LR130564 Australia 2018
1A
1B
2A
2B
LS992185 Germany 2018
95
LM997134 Norway 2010
CP024056 Canada 2004
CP024127 China 2014
AP010960 Japan 2008
LR134080 United Kingdom 2018
LR134204 Citrobacter koseri United Kingdom 2018
0.06 0.05 0.04 0.03 0.02 0.01 0.00
Gambar 2. Analisis filogenetik dengan metode UPGMA sekuens gen iutA asal Tabanan (MK776781) dan Badung (MK776782) dengan data gen iutA dari berbagai negara di dunia. Tanda lingkaran hitam menandakan gen iutA Escherichia coli asal Bali.
Semua data pada MEGA lalu dibuat situs polimorfik (polymorphic sites). Hasil analisis menunjukkan 43 situs basa yang berbeda di tingkat asam nukleat (tabel 1). Pada tingkat asam amino terdapat 13 situs polimorfik (tabel 2). Berdasarkan hasil tersebut, Substitusi T108C dan T192C unik untuk klaster isolat dalam studi ini bersama
isolat KP657535 Brazilia 2011, LT599825 Jerman 2016, dan CP033635 Cina 2016. Substitusi itu termasuk mutasi bisu (silent mutation), yaitu tidak menyebabkan perubahan asam amino.
Pembahasan
Gen iutA merupakan gen patogen pada strain APEC yang merupakan reseptor untuk ferric aerobactin dalam proses penyadapan zat besi pada protein sel inang (Rodriguez-Siek et al., 2005). Gen ini ditemukan dalam persentase yang cukup tinggi dari isolat kolibasilosis unggas di berbagai negara, seperti di Amerika Serikat sebesar 79,5% (Johnson et al., 2006), di Zimbabwe sebesar 80% (Mbangai dan Nyararai, 2015), di Cina sebesar 70,3% (Wang et al., 2015), dan di Eropa (Prancis, Spanyol, dan Belgia) sebesar 82,7% (Schouler et al., 2012). Frekuensi gen ini lebih tinggi pada isolat patogen, yaitu sekitar 79,5%, dibandingkan dengan bakteri komensal, yaitu sekitar 34,2% (Johnson et al., 2006).
Dalam penelitian ini, gen iutA berhasil diamplifikasi dengan spesifik
menggunakan sepasang DNA primer gen iutA yang sudah dipublikasi (Johnson et al., 2006). Hasil berupa pita DNA tunggal dengan panjang sekitar 302 bp. Kemudian hasil sekuensing DNA gen iutA yang dapat dianalisis dengan baik yaitu sepanjang 250 bp.
Sekuens gen iutA sampel MK776781 dari kabupaten Tabanan dan MK776782 dari kabupaten Badung tidak memiliki perbedaan urutan basa DNA. Hal tersebut membuktikan bahwa gen iutA APEC pada ayam buras asal Tabanan dan Badung memiliki homologi DNA 100%. Hal ini dapat terjadi karena APEC yang bersirkulasi di Bali berasal dari satu asal-usul yang sama. Disamping itu, sekuen DNA makhluk hidup cenderung stabil. Tingkat mutasi DNA E. coli diperkirakan 2-8 × 10-4 (Kibota and Lynch, 1996; Boe et al., 2000).
Tabel 1. Situs polimorfik gen iutA yang dideteksi dari Escherichia coli asal Bali (MK776781 dan MK776782) dengan data yang tersedia di genbank di tingkat asam nukleat
Urutan Basa
Geneid, Negara, Tahun K
1111111111111111111222222
112234566778899990012335567888899999013444
5561495239781403785800280601016823568438235
LC056228 India 2015
CP021179 United Emirates
Arab 2009
CP018994 Israel 2012
CP019018 Argentina 2010
CP006784 Denmark 2013
CP010372 Colombia 2012
LM996880 Norway 2010 CP009579 Netherlands 2014
CP023826 Sweden 2009
MH753063 Malaysia 2018
MK111112 Iran 2012
CP022731 Saudi Arabia 2012
KP119165 Canada 2014
CP031653 UK London 2016
MH422552 Spain 2015
AP018784 Japan 2007
CP026643 South Korea 2017
CP035516 USA 2015
CP035377 Brazil 2015
LR130556 Australia 2018
CP035124 China 2017
CP031295 Vietnam 2017
CP027851 Mexico 2010
LS992168 Germany 2018
CP025403 Qatar 2016
CP019055 Hongkong 2014 CP022069 S.enterica USA 2011
KY293686 Russia 2016
CP023145 Egypt 2016
CP023363 UK Scotland 2002
CP024827 South Korea
2015
CP029978 Switzerland 2018
CP030791 USA 1982
MG649062 Italy 2017
CP028304 Pakistan 2016
1A GGTCACCGTCGCCTCAACGATCCTTGTGTGCCTCGGAGTTGAT
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
1A
1A
1A ...........................................
1A ...........................................
1A ...........................................
1A ...........................................
CP026854 Slovakia 2018
CP027119 France 2010
MK776782 INDONESIA
(BALI) 2018
MK776781 INDONESIA
(BALI) 2018
KP657535 Brazil 2011
LT599825 Germany 2016
CP033635 China 2016
1A
1A
1B
1B
1B
1B
1B
|
........................................... | |
|
................... C...... |
...... C.......... |
|
................... C...... |
...... C.......... |
|
................... C...... |
...... C.......... |
|
................... C...... |
...... C.......... |
|
CP024992 Germany 2011 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CP016828 South Korea 2008 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CP011331 Denmark 2011 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CP025707 USA 2018 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CU928145 France 2008 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CP034067 Shigella sonnei USA 2015 |
2A |
. ACAGTAA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. . . A. . |
. GA |
|
CP024056 Canada 2004 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CP024127 China 2014 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CP021175 Taiwan 2016 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CP014092 USA 2018 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CU928163 France 2008 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
LM997134 Norway 2010 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
AP010960 Japan 2008 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CP012693 South Korea 2003 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
LR130564 Australia 2018 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
CP025573 Sweden 2008 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
LS992185 Germany 2018 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
|
LR134080 United Kingdom 2018 |
2B |
. ACAGTGA. . . |
. TC. C. GA. AGTC..... |
. TAC. |
. A. A. . |
. G. |
LR134204 Citrobacter koseri United Kingdom 2018
Outg
roup A. C. G. . . GTAT. CTCG. . . AG. . CTCACA. ACTT. T. CCTG.
Keterangan: K: Kelompok; Data diseleksi dari genbank Gen iutA dari kode akses MK776781 dan kode akses MK776782 berada dalam kelompok 1B
Tabel 2. Situs polimorfik gen iutA yang dideteksi dari Escherichia coli asal Bali (A2981 dan
A2982) dengan data yang tersedia di genbank di tingkat asam amino
Urutasn Situs Asam Amino
|
Geneid, Negara, Tahun |
K |
1 2 8 |
2 6 |
3 1 |
3 3 |
3 7 |
4 0 |
6 1 |
6 3 |
6 6 5 6 |
8 1 |
8 2 |
|
LC056228 India 2015 CP021179 United Emirates Arab 2009 CP018994 Israel 2012 CP019018 Argentina 2010 CP006784 Denmark 2013 CP010372 Colombia 2012 LM996880 Norway 2010 CP009579 Netherlands 2014 |
1A 1A 1A 1A 1A 1A 1A 1A |
R D G R T F N D A L G R F | ||||||||||
|
CP023826 Sweden 2009 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
MH753063 Malaysia 2018 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
MK111112 Iran 2012 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP022731 Saudi Arabia 2012 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
KP119165 Canada 2014 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP031653 UK London 2016 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
MH422552 Spain 2015 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
AP018784 Japan 2007 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP026643 South Korea 2017 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP035516 USA 2015 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP035377 Brazil 2015 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
LR130556 Australia 2018 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP035124 China 2017 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP031295 Vietnam 2017 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP027851 Mexico 2010 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
LS992168 Germany 2018 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP025403 Qatar 2016 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP019055 Hongkong 2014 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP022069 S.enterica USA 2011 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
KY293686 Russia 2016 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP023145 Egypt 2016 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP023363 UK Scotland 2002 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP024827 South Korea 2015 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP029978 Switzerland 2018 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP030791 USA 1982 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
MG649062 Italy 2017 |
1A |
... |
.. |
... |
.. |
.. |
. |
|
CP028304 Pakistan 2016 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP026854 Slovakia 2018 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP027119 France 2010 |
1A |
. . . |
. . |
. . . |
. . |
. . |
. |
|
MK776782 INDONESIA (BALI) 2018 |
1B |
. . . |
. . |
. . . |
. . |
. . |
. |
|
MK776781 INDONESIA (BALI) 2018 |
1B |
. . . |
. . |
. . . |
. . |
. . |
. |
|
KP657535 Brazil 2011 |
1B |
. . . |
. . |
. . . |
. . |
. . |
. |
|
LT599825 Germany 2016 |
1B |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP033635 China 2016 |
1B |
. . . |
. . |
. . . |
. . |
. . |
. |
|
CP024992 Germany 2011 |
2A |
. N . |
S S |
Y K . |
D . |
. . |
Y |
|
CP016828 South Korea 2008 |
2A |
. N . |
S S |
Y K . |
D . |
. . |
Y |
|
CP011331 Denmark 2011 |
2A |
. N . |
S S |
Y K . |
D . |
. . |
Y |
|
CP025707 USA 2018 |
2A |
. N . |
S S |
Y K . |
D . |
. . |
Y |
|
CU928145 France 2008 |
2A |
. N . |
S S |
Y K . |
D . |
.. |
Y |
|
CP034067 Shigella sonnei USA 2015 |
2A |
. N . |
S S |
Y K . |
D . |
.. |
Y |
|
CP024056 Canada 2004 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CP024127 China 2014 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CP021175 Taiwan 2016 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CP014092 USA 2018 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CU928163 France 2008 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
LM997134 Norway 2010 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
AP010960 Japan 2008 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CP012693 South Korea 2003 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
LR130564 Australia 2018 |
2B |
. N . |
S S |
Y K . |
D . |
R . |
. |
|
CP025573 Sweden 2008 |
2B . |
N . |
S |
S Y K . D . |
R . . |
|
LS992185 Germany 2018 |
2B . |
N . |
S |
S Y K . D . |
R . . |
|
LR134080 United Kingdom 2018 |
2B . |
N . |
S |
S Y K . D . |
R . . |
|
LR134204 Citrobacter koseri United Kingdom 2018 |
Outgroup Q |
. D S |
A Y K N D F |
. L . | |
Keterangan: K: Kelompok; Data diseleksi dari genbank
Hasil analisis situs polimorfik gen iutA menunjukkan 43 situs polimorfik ditingkat asam nukleat dan 13 situs ditingkat asam amino dari 61 sekuens yang ada. Gen iutA asal Bali memiliki homologi 100% dengan gen iutA asal KP657535 Brazilia (2017), LT599825 Jerman (2016), dan CP033635 Cina (2016). Hal ini menunjukan kekerabatan gen iutA E. coli asal Bali dengan negara tersebut. Asal-usul gen iutA E. coli Bali sama dengan isolat dari tiga negara tersebut. Hal tersebut dapat terjadi karena DNA E. coli memiliki kemungkinan yang sangat kecil untuk mengalami mutasi
Penelitian tentang gen iutA harus dilakukan di seluruh provinsi di Indonesia untuk mengetahui frekuensi gen iutA pada APEC. Kajian lebih lanjut tentang gen iutA dan gen patogenik lainnya dapat membantu untuk pengembangan obat antimikrobial yang secara khusus menargetkan E. coli patogen atau APEC. Selain itu kajian tentang gen patogenik APEC dapat membantu untuk mendiagnosis secara cepat. Kajian lebih lanjut tentang sifat gen patogenik APEC dan kerja sistem imun unggas dapat membantu untuk pengembangan vaksin APEC yang akan sangat menguntungkan peternakan ayam.
Beberapa penelitian telah mengkaji tentang vaksin kolibasilosis unggas, seperti iss-based vaccine yang berpotensi menargetkan tiga serotipe APEC yaitu O2, 078 dan O1 (Lynne et al., 2012). Vaksin Poulvac Escherichia coli yang menghilangkan gen aroA pada serotipe O78 (Uotani et al., 2017). Gen iutA memiliki potensi yang cukup besar untuk dikembangkan sebagai vaksin APEC. Hal ini dikarenakan persentase gen iutA pada isolat APEC cukup tinggi di berbagai negara.
SIMPULAN DAN SARAN
Simpulan
Gen iutA dapat dideteksi dari dua isolat Avian Pathogenic Escherichia coli (APEC) asal ayam buras di Tabanan dan Badung tahun 2018. Kedua isolat tersebut mempunyai gen iutA dengan sekuen yang sama (homologi 100%). Hasil analisis filogenik gen iutA asal Tabanan dan Badung dengan gen iutA dari seluruh negara menunjukan kekerabatan dengan gen iutA isolat E. coli asal Brazilia (KP657535) tahun 2011, Jerman (LT599825) tahun 2016, dan Cina (CP033635) tahun 2016.
Saran
Penelitian dan pendataan perlu dilakukan di seluruh provinsi di Indonesia tentang gen iutA dan gen patogenik APEC lainnya. Selain itu, uji biologis perlu juga dilakukan untuk mengetahui dampak gen iutA pada sifat patogenik E.coli.
UCAPAN TERIMAKASIH
Ucapan terimakasih ditujukan kepada drh. Ni Made Ritha Krisna Dewi, S.KH, M.Si dan drh. Ni putu Sutrisna Dewi, S.KH selaku asisten laboratorium yang telah membantu dalam penelitian ini. Penelitian ini didanai oleh The Professor and Promotion Project, Universitas Udayana, Bali, DIPA-PNBP 2018, nomor kontrak 383-1/UN14.4.4.A/LT/2018, tertanggal 28 maret 2018.
DAFTAR PUSTAKA
Abalaka SE, Sani NA, Idoko IS, Tenuche OZ, Oyelowo FO, Ejeh SA, Enem SI. 2017 Pathological changes associated with an outbreak of colibacillosis in a
commercial broiler flock. Sokoto J. Vet. Sci. 15(3): 95-102.
Barus DO, Gelgel KTP, Suarjana IGK. 2013. Uji kepekaan bakteri escherichia coli asal ayam pedaging terhadap antibiotik doksisiklin, gentamisin, dan tiamfenikol. Indonesia Medicus
Veterinus. 2(5): 538-545.
Brown TA. 1992. Genetics: Molecular Approach, Second Edition. Chapman & Hall, London.
Boe L, Danielsen M, Knudsena S, Petersen JB, Maymann J, Jensen PR. 2000. The frequency of mutators in populations of Escherichia coli. Mutation Res. Fund. Mol. Mechanisms Mutagenesis. 448(1): 47–55.
Guabiraba R, Schouler C. 2015. Avian colibacillosis: still many black holes. FEMS Microbiol. Letters. 362(12): 1-8.
Handoyo D, Rudiretna A. 2000. Prinsip umum dan pelaksanaan polymerase chain reaction (PCR). Unitas, 9(1):17-29
Johnson TJ, Siek KE, Johnson SJ, Nolan LK. 2006. DNA sequence of a coiv plasmid and prevalence of selected plasmid-encoded virulence genes among avian escherichia coli strains. J. Bacteriol. 188(2): 745-758.
Johnson TJ, Wannemuehler Y, Doetkott C, Johnson SJ, Rosenberg SC, Nolan LK. 2008. Identification of minumal predictors of avian pathogenic escherichia coli virulence for use as a rapid diagnostic tool. J. Clin.
Microbiol. 46(12): 3987-3996.
Kabir SML. 2010. Avian colibacillosis and salmonellosis: a closer look at
epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Pub. Health. 7(1): 89114.
Kibota TT, Lynch M. 1996. Estimate of the genomic mutation rate deleterious to overall fitness in E. coli. Nature. 381(6594): 694–696.
Landman WJMM, van Eck JHH. 2016. The incidence and economic impact of the Escherichia coli peritonitis syndrome in
Dutch poultry farming. Avian Pathol. 44(5): 370-378.
Lau GL, Sieo CC, Tan WS, Ho YW. 2012. Characteristic of phage effective for colibacillosis control in poultry. J. Sci. Food Agric. 92(13): 57-63.
Luhung YGA, Suarjana IGK, Gelgel KTP. 2017. Sensitivitas isolat escherichia coli patogen dari organ ayam pedagng terinfeksi koliseptisemia terhadap oksitetrasiklin, ampisilin dan
sulfametoksazol. Buletin Veteriner Udayana. 9(1): 60-66.
Lynne AM, Kariyawasam S,
Wannemuehler Y, Johnson TJ, Johnson SJ, Sinha AS, Lynner DK, Moon HW, Jordan DM, Logue CM, Foley SL, Nolan LK. 2012. Recombinant iss as a potential vaccine for avian colibacillosis. Avian Dis. 56(1): 192199.
Rodriguez-Siek KE, Giddings CW, Doetkott C, Johnson TJ, Nolan LK. 2005. Characterizing the APEC pathotype. Vet. Res. 36(2): 241–256.
Schouler C, Schaeffer B, Brée A, Mora A, Dahbi G, Biet F. 2012. Diagnostic strategy for identifying avian pathogenic escherichia coli based on four patterns of virulence genes. J. Clin. Microbiol. 50(5): 1673–1678.
Tabrah FL. 2011. Koch’s postulates, carnivorous cows, and tuberculosis today. Hawai‘i Med. J. 70(7): 144-148.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28(10): 2731-2739.
Tarmudji. 2003. Kolibasiolosis pada ayam: etiologi, patologi dan pengendaliannya. Wartazoa 13(2):65-73.
Uotani Y, Kitahara R, Imai T, Tsutsumi N, Sasakawa C, Nagai S, Nagano T. 2017. Efficacy of an avian colibacillosis live vaccine for layer breeder in Japan. J. Vet. Med. Sci. 79(7): 1215–1219.
Walsh SP, Metzger DA, Higuchi R. 1991.
Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques. 10(4): 506-513.
Wang J, Tang P, Tan D, Wang L, Zhang S, Qiu Y, Dong R, Liu W, Huang J, Chen T, Ren J, Lil C, Liu H. 2015. The pathogenicity of chicken pathogenic
escherichia coli is associated with the numbers and combination patterns of virulence-associated genes. J. Vet. Med. 5: 243-254.
Whittam TS, Wilson RA. 1988. Genetic relationships among pathogenic strains of avian Escherichia coli. Infect. Immun. 56(9): 2458–2466.
66
Discussion and feedback