Early study of the flowering and fruiting periods of Sauromatum horsfieldii Miq. (Araceae) in Bali
on
JURNAL BIOLOGI UDAYANA 26(1): 1-10
P ISSN: 1410-5292 E ISSN: 2599-2856
Studi awal masa berbunga dan berbuah Sauromatum horsfieldii Miq. (Araceae) di Pulau Bali
Early study of the flowering and fruiting periods of Sauromatum horsfieldii Miq. (Araceae) in Bali
Agung Kurniawan1,2 *, Sin Yeng Wong3
-
1)Pusat Penelitian Konservasi Tumbuhan dan Kebun Raya-LIPI, Indonesia – 16003
-
2)Balai Konservasi Tumbuhan, Kebun Raya “Eka Karya” Bali-LIPI, Indonesia – 82191
3)Institute of Biodiversity and Environmental Conservation, Universiti Malaysia Sarawak, Samarahan, Sarawak, Malaysia – 94300
*Email: aracasia@gmail.com
Diterima 14 Juli 2021 Disetujui xx xxx xx
INTISARI
Studi masa berbunga dan berbuah Sauromatum horsfieldii Miq. (Araceae) di habitat alaminya adalah observasi biologi bunga Araceae pertama yang dilakukan di Bali. Studi awal ini bertujuan untuk mengetahui masa berbunga dan berbuah S. horsfieldii yang hidup secara alami di area Kebun Raya “Eka Karya” Bali - LIPI. Hasil observasi memperlihatkan bahwa durasi berbunga S. horsfieldii memerlukan waktu >16 hari, sementara durasi berbuah memakan waktu ±60 hari. Satu siklus masa matang (anthesis) bunga betina dan jantan berlangsung selama ±72 jam. Bunga betina lebih dahulu mengalami anthesis dibandingkan dengan bunga jantan. Serangga diduga merupakan hewan penyerbuk yang membantu keberhasilan proses penyerbukan bunga.
Kata kunci: anthesis, masa berbunga dan berbuah, Sauromatum horsfieldii
ABSTRACT
The study of flowering and fruiting periods of Sauromatum horsfieldii Miq. (Araceae) in its natural habitat is the first observation on the flowering biology of an aroid in Bali. This preliminary study aimed to investigate the flowering and fruiting behaviour of S. horsfieldii in its natural habitat at Bali Botanic Garden. The result showed that it took approximately more than 16 days for flowering period, while fruiting duration took about 60 days. Blooming (anthesis) occurred for approximately 72 hours. Female flowers matured prior to male flowers. Insects were proposed as pollinator which played a significant role to reproductive success.
Keywords: anthesis, flowering and fruiting periods, Sauromatum horsfieldii
PENDAHULUAN
Jenis Sauromatum horsfieldii Miq. tergolong ke dalam suku Araceae, yang memiliki karakter perbungaan uniseksual. Pada tipe perbungaan ini,
bunga jantan dan betina terpisah pada tempat yang berbeda dalam satu tongkol (spadix). Kelompok bunga jantan terletak pada spadix bagian atas, sedangkan kelompok bunga betina
berada pada spadix bagian bawah (Grayum, 1990; Mayo et al., 1997).
Suku Araceae merupakan suku yang protoginous, dimana masa matang (anthesis) bunga betina dan bunga jantan tidak terjadi dalam waktu yang bersamaan, sehingga tidak dapat menyerbuki sendiri. Umumnya proses penyerbukan dibantu oleh serangga, seperti: lalat, lebah, kepik, dan kumbang (Grayum, 1990; Mayo et al., 1997; Albre et al., 2003; Gibernau, 2003; Gibernau, 2011). Serangga penyerbuk tertarik oleh adanya emisi yang menyerupai bau kotoran, bangkai, ataupun daging busuk. Pada tribus Areae, dimana Sauromatum tergolong di dalamnya, bau busuk tersebut dihasilkan oleh appendix, bagian bunga jantan steril yang terletak di ujung spadix (Grayum, 1990; Mayo et al., 1997; Hentrich et al., 2010; Boyce & Wong, 2012) dan bunga steril yang berbentuk gada yang terletak di tengah spadix (Hadacek & Weber, 2008). Hal lain yang berkaitan dengan proses anthesis adalah produksi panas (thermogenesis). Thermogenesis merupakan proses meningkatnya suhu bunga dibanding suhu di sekitarnya, yang ditandai dengan perubahan fisiologis pada bunga seperti: stigma menjadi reseptif dan diproduksinya bau (Meeuse, 1975; Meeuse, 1978; Grayum, 1990; Mayo et al., 1997; Seymour & Schulze-Motel, 1997; Seymour, 2001; Gibernau & Barabe, 2002).
Beberapa studi terkait biologi bunga pada marga Sauromatum, umumnya dilakukan pada jenis Sauromatum gutattum Schott (Meeuse & Hatch, 1960; Meeuse, 1978; Skubatz et al., 1991; Skubatz et al., 1996). Belum ada publikasi penelitian biologi bunga yang dikerjakan pada jenis S. horsfieldii pada habitat alami khususnya di Bali, bahkan di Indonesia. Adapun penelitian S. horsfieldii yang dilakukan Yuzammi et al. (2017) adalah tentang gambaran umum, konservasi dan metode perbanyakannya. Oleh karena itu, kegiatan studi awal biologi bunga S. horsfieldii ini dilakukan untuk mengetahui masa berbunga dan berbuah di habitat aslinya, yaitu di kawasan Kebun Raya “Eka Karya” Bali. Data yang diperoleh diharapkan dapat menjadi data dasar
yang dapat dimanfaatkan untuk penelitian tahap berikutnya, seperti studi fitokimia tentang kandungan emisi ataupun studi penyerbukan dan keberhasilan reproduksi.
BAHAN DAN METODE
Tempat penelitian
Observasi dilakukan di dalam kawasan Kebun Raya “Eka Karya” Bali-LIPI (KRE) yang berdekatan dengan Cagar Alam (CA) Batukaru (Gambar 1). Kawasan ini berada di daerah pegunungan dengan ketinggian 1250-1400 m dpl., kisaran suhu udara antara 14-22,5oC, kelembaban udara 70-96%, intensitas cahaya matahari 45-60%, dan curah hujan 2000-3000 mm/tahun (Putri, 2018; https://krbali.lipi.go.id/sejarah.html). Kondisi area observasi berada di bawah tegakan tanaman reboisasi rasamala, Altingia excelsa Noronha (Altingiaceae) hasil penanaman tahun 1958 (Siregar & Undaharta, 2014), yang berfungsi sebagai batas area sekaligus zona penyangga hutan alam CA Batukaru (Siregar & Undaharta, 2018). Area ini dipenuhi dengan seresah yang didominasi oleh dedaunan kering A. excelsa, dengan tebal lebih kurang 5-10 cm. Pada umumnya area petak koleksi KRE selalu mendapatkan pemeliharaan berupa pemangkasan gulma, namun di beberapa lokasi termasuk di area observasi, hal ini tidak terjadi ataupun jarang dilakukan karena hampir tidak ada rumput atau gulma lain yang tumbuh kecuali beberapa benih pohon atau tumbuhan paku liar. Hal tersebut diharapkan menjadikan kegiatan observasi ini tidak terganggu. Kegiatan ini dilakukan pada tahun 2016, pada rentang waktu bulan Februari -April bertepatan dengan puncak keberadaan populasi S. horsfieldii di musim hujan. Secara umum, jangka waktu berbunga dan berbuah S. horsfieldii dimulai pada bulan Desember hingga Mei di lokasi observasi. Periode tersebut akan berbeda di tempat lain, seperti di India terjadi pada bulan April-Juli (Odyuo et al., 2015).
Objek penelitian yang digunakan adalah tumbuhan liar S. horsfieldii yang terdapat di KRE. Jenis ini pertama kali teridentifikasi sebagai S. horsfieldii di area KRE pada tahun 2010 oleh P.C. Boyce dan dipublikasikan dalam Flora of Thailand (Boyce et al., 2012). Selain di Bali, jenis ini ditemukan juga di Jawa, Sumatra, Semenanjung Malaysia, Thailand, Vietnam, Myanmar, Kamboja, Laos, China, dan India (Heng & Hetterscheid, 2010; Boyce et al., 2012; Odyuo et al., 2015; Yuzammi et al., 2017). Berdasarkan pengamatan visual sebelumnya (pra-observasi), jenis ini hanya tumbuh di bagian barat kawasan KRE yang berbatasan dengan lokasi CA Batukaru di lereng bagian timur Gunung Tapak (Gambar 1). Tumbuhan ini banyak ditemukan pada musim hujan di area ini, sebaliknya jarang sekali atau bahkan tidak ditemukan pada saat musim kemarau karena umbinya mengalami dormansi.
Gambaran Umum S. horsfieldii (Gambar 2)
Jenis Sauromatum horsfieldii merupakan tumbuhan herba dengan tinggi 50-60 cm, memiliki umbi yang berukuran relatif kecil dengan diameter kurang dari 10 cm, berbentuk bulat pipih dan berwarna coklat muda hingga krem, bagian dalam umbi berwarna putih-krem (Gambar 2a). Tangkai daun berwarna hijau dengan variasi warna kecoklatan berupa strip atau bergaris pendek. Daun menjari berbentuk seperti tapak kaki (pedate), berwarna hijau muda hingga hijau gelap dan mengkilat pada bagian atas. Anak daun berbentuk lonjong atau memanjang, tepi daun rata, dan ujung runcing (Gambar 2b). Perbungaan terdiri dari seludang dan tongkol (spadix). Seludang bagian atas berbentuk silinder yang mengerucut di ujung atas, berwarna hijau
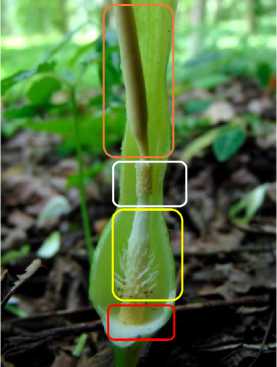
muda hingga putih kekuningan (pada saat anthesis bunga jantan), dengan ukuran lebih panjang dibandingkan seludang bagian bawah. Seludang bagian bawah berbentuk bulat telur dan berwarna hijau hingga hijau tua (Gambar 2c dan 2d). Di bagian tongkol melekat beragam tipe bunga. Kelompok bunga jantan fertil berada di tongkol bagian atas dan tertutup seludang bagian atas. Di sebelah atas zona bunga jantan fertil terdapat appendiks berbentuk silindris berukuran hampir separuh panjang tongkol. Kelompok bunga betina fertil berada di tongkol bagian bawah dan tertutup seludang bagian bawah. Di antara keduanya terdapat kelompok bunga steril yang bermodifikasi bentuk (Gambar 2e). Buah S. horsfieldii tergolong buah beri berbentuk bulat hingga bulat lonjong, kulit buah berwarna hijau pucat hingga hijau saat masak yang berisi berwarna putih dengan ujung kehijauan. Biji berwarna putih kehijauan (Gambar 2e dan 2f).
Pengumpulan Data
Kegiatan observasi lapangan S. horsfieldii diawali dengan menelusuri keberadaan populasinya di area KRE, memeriksa kondisi morfologinya apakah dalam keadaan berbunga atau tidak, menentukan individu dewasa yang akan berbunga (memiliki kuncup perbungaan), serta mengamati periode berbunga dan berbuah yang dilakukan pada siang dan malam hari sesuai dengan tahapan perkembangan dan mendokumentasikannya dengan menggunakan kamera. Waktu pengamatan ini dipilih karena diduga jenis ini mengalami masa anthesis lebih dari 24 jam, seperti halnya beberapa jenis Araceae lainnya (Skubatz et al., 1991; Barabe et al., 2002; Albre et al., 2003).

Gambar 1. Peta lokasi observasi pada peta Kebun Raya “Eka Karya” Bali (Sumber: Unit Registrasi, Kebun Raya “Eka Karya” Bali, 2015). Lokasi observasi ditunjukan oleh anak panah berwarna putih. Sebaran populasi Sauromatum horsfieldii Miq. ditunjukkan oleh lingkaran berwarna kuning. Lokasi S.horsfieldii yang baru ditemukan pada tahun 2021 ditandai oleh lingkaran berwarna merah.


(b)

(c)

(d)
(e)

(f)
Gambar 2. Perawakan Sauromatum horsfieldii Miq. (a) Umbi; (b) Daun; (c) Anakan dan kuncup perbungaan; (d) Perbungaan dewasa; (e) Zona bunga jantan (kotak putih), zona betina (kotak merah), zona bunga steril (kotak kuning), dan appendiks (kotak oranye); (f) Buah dan biji.
HASIL
Berdasarkan hasil observasi terhadap sebaran populasi S. horsfieldii di KRE, menunjukkan bahwa jenis ini paling banyak ditemukan di bagian barat KRE yang berbatasan dengan CA Batukaru bagian timur, di lereng Gunung Tapak (Gambar 1). Di dalam area tersebut dapat ditemukan lebih dari 50 individu S. horsfieldii. Jenis ini juga dapat ditemukan di beberapa bagian lain di dalam KRE, namun demikian jumlah individunya lebih sedikit.
Beberapa tahap proses pembentukan bunga S. horsfieldii dari satu individu yang dilakukan selama 16 hari di bulan Maret 2016 ditunjukkan pada Gambar 3. Tahap yang pertama dimulai dari kuncup perbungaan (Gambar 3a), kemudian
terlihat celah atau lubang kecil di bagian seludang bagian bawah (Gambar 3b). Seludang bagian atas mulai terbuka pada keesokan harinya (Gambar 3c-3d) dan terlihat serbuk sari yang jatuh dan menempel pada seludang bagian dalam pada hari kedua setelah seludang bagian atas terbuka (Gambar 3e). Selanjutnya warna seludang atas semakin memudar, menguning, dan menutup kembali, lambat laun menjadi layu (Gambar 3f-3g). Seludang atas akhirnya membusuk dan berwarna kuning sedangkan seludang bawah menjadi lebih gelap warnanya (Gambar 3h). Perbungaan ini selanjutnya memasuki tahap pembentukan buah apabila penyerbukan berhasil. Data waktu observasi pada Gambar 3 menunjukkan bahwa periode berbunga S. horsfieldii membutuhkan waktu lebih dari 16 hari.

(a)

(b)

(c) (d)

(e)

(f)

(g) (h)
Gambar 3. Tahap pembentukan bunga S. horsfieldii. (a) Kuncup perbungaan, hari ke-1 observasi (14 Maret 2016); (b) Kuncup perbungaan yang siap mekar, dengan celah/lubang kecil pada seludang bawah (lingkaran putih), hari ke-7 (21 Maret 2016); (c) Seludang atas mekar tampak samping, hari ke-8 (22 Maret 2016), dengan celah kecil masih terbuka; (d) Seludang atas mekar tampak depan, hari ke-8 (22 Maret 2016); (e). Serbuk sari jatuh dari bunga jantan, hari ke-9 (23 Maret 2016, dini hari); (f) Seludang atas mulai layu dan warna memucat, hari ke-11 (25 Maret 2016, dini hari); (g) Seludang atas layu dan mengatup kembali, hari ke-13 (27 Maret 2016); (h) Seludang atas layu, kuning, seludang bawah hijau tua, hari ke-16 (30 Maret 2016).
Tahap pembentukan buah pada individu yang berbeda dengan Gambar 3, diperlihatkan pada Gambar 4. Observasi dilakukan pada perbungaan dengan seludang bunga yang sudah dikelupas sehingga proses pembentukan buah dapat lebih terlihat dengan jelas. Secara umum, perubahan dari hari ke hari cenderung lambat, sehingga gambar yang disajikan dibuat dalam rentang waktu ±7 hari. Setelah terjadi penyerbukan (Gambar 4a), pengamatan tidak dilakukan selama 2 minggu, hingga wujud buah sudah terbentuk (Gambar 4b). Semakin lama, buah semakin membesar dan kulit buah berwarna hijau gelap (Gambar 4c-4g). Pada akhirnya kulit buah terbelah, sebagai penanda buah telah masak dan kulit buah berubah warna menjadi hijau pucat
(Gambar 4h). Namun demikian keesokan harinya tepat setelah buah berumur 2 bulan atau 60 hari, buah tersebut hilang atau mungkin dimakan hewan liar.
PEMBAHASAN
Seperti halnya tipe Araceae dengan tipe life forms geofit yang dicirikan dengan adanya umbi, tumbuhan ini mengalami masa dormansi pada saat musim kemarau atau dorman musiman (Mayo et al., 1997; Boyce & Wong, 2012). Hasil observasi menunjukkan, pada saat musim hujan, S. horsfieldii mengalami pecah dormansi dan banyak ditemukan secara mengelompok di area observasi. Anakan S. horsfieldii yang ditemukan
satu atau dua individu diduga berasal dari umbi yang pecah dormansi, sedangkan anakan yang ditemukan secara bergerombol dalam satu titik diduga berasal dari biji. Meskipun demikian,
masih belum banyak diketahui viabilitas biji Sauromatum (Mayo et al., 1997), sehingga dugaan awal pemencaran biji tersebut perlu dilakukan studi lebih dalam.

(a) (b) (c) (d)

(e) (f) (g)
Gambar 4. Tahap pembentukan buah S.horsfieldii. (a) Hari ke-1 bunga yang sudah terjadi penyerbukan (1 Maret 2016); (b) Buah muda sudah terbentuk, hari ke-14 (14 Maret 2016); (c) Buah, hari ke-21 (21 Maret 2016); (d) Hari ke-29 (29 Maret 2016); (e) Hari ke-36 (4 April 2016); (f) Hari ke-43 (11 April 2016); (g) Hari ke-50 (18 April 2016); (h) Buah masak, hari ke-59 (27 April 2016).

(h)
Gambar 3b memperlihatkan bahwa bunga betina sudah memasuki tahap anthesis pada hari ke-7 yang ditandai adanya celah atau lubang kecil pada seludang bagian bawah. Namun belum teramati sejak kapan celah kecil tersebut mulai terbentuk. Keesokan harinya (hari ke-8) seludang bagian atas mulai terbuka yang menunjukkan bahwa bunga jantan memasuki masa anthesis (Gambar 3c) dan menjadi akhir masa anthesis bunga betina. Pada hari ke-9, bunga jantan memasuki puncak anthesis yang dicirikan dengan luruhnya serbuk sari terjadi saat dini hari antara pukul 03.00-04.00 WITA (Gambar 3e). Selanjutnya pada hari ke-11 seludang atas layu yang menunjukkan akhir masa anthesis bunga jantan (Gambar 3f). Secara singkat, hal ini menggambarkan bahwa masa matang bunga tidak
terjadi secara bersamaan (protoginous) melainkan masa matang bunga betina terjadi lebih dahulu dibandingkan bunga jantan. Dalam satu siklus berbunga (anthesis bunga jantan dan betina), durasi waktu yang diperlukan cukup singkat yaitu ±72 jam atau 3 hari. Namun demikian tidak regulernya rentang waktu pengamatan menyebabkan belum diketahui secara persis berapa lama total waktu berbunga dari awal anthesis hingga akhir masa berbunga. Sebagai acuan, pada S. guttatum masa anthesis bunga betina berlangsung lebih dulu setelah tengah malam hingga dini hari lalu diikuti oleh masa anthesis bunga jantan (Meeuse & Buggeln, 1969; Buggeln et al., 1971; Dakwale & Bhatnagar, 1985). Selain itu belum diketahui kondisi reseptif kepala putik bunga betina karena observasi
dilakukan dengan kondisi seludang bawah yang tertutup. Sebuah lubang kecil (jendela) perlu dibuat pada seludang bawah untuk mengamati kondisi bunga betina pada masa anthesis.
Sauromatum horsfieldii memiliki sekelompok organ unik di bagian spadix yang terletak di zona steril (Gambar 2e). Zona bunga steril yang memisahkan zona bunga betina dan zona bunga jantan menjadi salah satu ciri utama dari tribus Areae (Boyce & Wong, 2012). Organ unik ini disebut staminodes atau bunga jantan steril yang memiliki dua tipe yaitu berbentuk gada dan silinder melengkung ke arah atas (Gambar 2e dan lingkaran merah pada Gambar 4a) (Boyce et al., 2012; Odyuo et al., 2015) dan kedua bentuk staminodes ini menjadi penentu karakter antara Sauromatum dengan Typhonium (Boyce & Wong, 2015). Staminodes ini berfungsi sebagai penghalang bagi serangga penyerbuk untuk melepaskan diri dari seludang bawah. Serangga penyerbuk akan memasuki celah kecil pada seludang bawah (Gambar 2b), terperangkap di dalam ruangan yang berisi bunga betina. Celah kecil ini tetap terlihat dari bagian luar seludang bawah meskipun masa anthesis bunga betina sudah selesai. Namun belum diketahui apakah celah ini tertutup dari bagian dalam ataukah terbuka.
Penyebab datangnya calon serangga penyerbuk ke bunga Sauromatum dan bagaimana mekanismenya dilaporkan secara mendalam pada S. guttatum oleh Meeuse & Hatch (1960), Dakwale & Bhatnagar (1985), Skubatz et al. (1996) dan McDougal et al. (2016). Walaupun dalam observasi ini tidak berhasil menemukan atau bahkan menangkap serangga penyerbuk, mekanisme penyerbukan yang serupa dengan S. guttatum diduga juga dapat terjadi pada S. horsfieldii yaitu serangga memasuki seludang bawah melalui celah kecil pada saat seludang atas masih tertutup (masa anthesis bunga betina) atau melalui seludang atas pada saat terbuka (masa anthesis bunga jantan). Apabila serangga tersebut membawa serbuk sari dari bunga jantan maka diharapkan penyerbukan bunga betina dapat terjadi segera setelah serangga tersebut
terperangkap di dalam seludang bawah. Apabila serangga tidak membawa serbuk sari, maka serangga tersebut akan tetap terperangkap di dalam seludang bawah hingga terlepas pada saat seludang atas terbuka dan serbuk sari jatuh. Dugaan ini masih memerlukan penguat yaitu dengan dilakukannya studi lanjutan tentang polinasi S. horsfieldii. Menurut Grayum (1990), kelompok serangga yang biasanya membantu penyerbukan perbungaan dengan karakter yang dimiliki S. horsfieldii, adalah kumbang dan lalat. Gibernau (2003) dan Gibernau (2011) berhasil menghimpun kelompok serangga penyerbuk bunga Sauromatum, dari kategori kumbang yaitu: Scarabaeidae dan Bruchidae; dari kategori lalat, yaitu: Sepsidae, Muscidae, Otitidae,
Sarcophagidae, Calliphoridae, Diptera.
Keberhasilan penyerbukan oleh serangga akan menghasilkan buah dalam jumlah banyak yang terbungkus dalam seludang bagian bawah S. horsfieldii. Buah yang terbentuk dapat diobservasi dengan mengelupas seludang bawah. Proses pembentukan buah memerlukan waktu yang panjang terutama pada tahap awal proses setelah berhasilnya penyerbukan. Secara visual, tidak ada perubahan yang signifikan dari pembentukan buah yang terjadi selama lebih kurang 60 hari atau 2 bulan, kecuali ukuran buah yang bertambah besar (Gambar 4b-4g) dan pecahnya kulit buah saat buah masak (Gambar 4h). Perubahan warna kulit buah hampir tidak terlihat secara jelas yaitu dari hijau gelap saat buah muda hingga hijau pucat saat buah masak.
Pada observasi lapangan ditemui kendala utama yaitu hilangnya objek pengamatan. Individu S. horsfieldii pada Gambar 3, tidak ditemukan lagi di lokasi penelitian pada hari ke-18 setelah masa berbunga berakhir. Diduga kuat, objek pengamatan dirusak atau mungkin umbi S. horsfieldii dimakan oleh satwa liar. Area hutan alam CA Batukaru yang bersebelahan dengan lokasi observasi, merupakan habitat berbagai satwa liar. Menurut BKSDA Bali (2021) (https://www.ksda-bali.go.id/ca-batukahu/), satwa liar berpotensi yang berada di kawasan CA Batukaru hanya sejenis kijang, Muntiacus
muntjak (Zimmerman, 1780). Namun demikian, fakta di lokasi observasi menunjukkan bahwa satwa liar lain juga ditemukan seperti monyet ekor panjang, Macaca fascicularis Raffles, 1821. Satwa liar ini kadang-kadang masuk hingga ke area KRE sejauh 500 - 1000 m (Gambar 1). Di samping itu, menurut beberapa petugas pemelihara koleksi KRE (komunikasi pribadi), satwa liar lain sejenis tikus hutan juga diduga memakan umbi dari jenis beberapa Araceae, termasuk jenis yang diamati.
Pada tahun 2021, S. horsfieldii dapat ditemukan di area yang berbeda, dimana di lokasi tersebut tidak ditemukan sebelumnya. Hal ini diperkirakan terjadi karena adanya banjir yang melanda area KRE pada bulan Desember 2016 dan awal tahun 2017. Limpasan air diduga membawa umbi dan tanaman S. horsfiedii ke sebelah Timur KRE menuju area yang lebih rendah (Gambar 1).
SIMPULAN
Simpulan yang diperoleh dari studi awal ini adalah bahwa durasi berbunga dan berbuah, dimulai kuncup perbungaan hingga buah masak, memerlukan waktu selama >76 hari. Periode ini terbagi menjadi >16 hari masa berbunga dan ±60 hari masa berbuah. Dalam satu siklus masa matang (anthesis) bunga jantan dan betina memakan waktu sekitar 72 jam atau 3 hari. Studi ini merupakan langkah awal bagi studi biologi bunga S. horsfieldii secara lebih mendalam. Dengan diketahui data awal masa berbunga dan berbuah jenis ini, termasuk di dalamnya karakter khusus perbungaan selama masa anthesis diharapkan penelitian lanjutan bisa dilakukan. Contoh: studi penyerbukan dan serangga polinatornya, studi thermogenesis dan emisi bunga pada saat anthesis, serta studi keberhasilan reproduksi.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Bapak Farid Kuswantoro, S.Si. atas diskusi yang bermanfaat tentang serangga penyerbuk. Penulis juga menyampaikan terima kasih kepada Prof.
Hanna Skubatz dari NeoPro Labs. atas kesediannya dalam memberikan salinan makalah penelitian tentang S. guttatum. Ucapan terima kasih juga ditujukan kepada Bapak I Made Sumerta, S.P., Bapak I Ketut Nukaya, Bapak I Wayan Armawa, dan staf Unit Registrasi KRE atas informasi seputar area studi dan penggunaan peta Kebun Raya “Eka Karya” Bali.
KEPUSTAKAAN
Albre J, Quilichini A, Gibernau M. 2003.
Pollination ecology of Arum italicum (Araceae). Bot. J. Linn. Soc. 141(2): 205– 214. DOI:10.1046/j.1095-
8339.2003.00139.x
Barabe D, Gibernau M, Forest F. 2002. Zonal thermogenetic dynamics of two species of Philodendron from two different subgenera (Araceae). Bot. J. Linn. Soc. 139(1): 79-86. DOI: 10.1046/j.1095-8339.2002.00040.x
BKSDA Bali. 2021. Cagar Alam Batukahu. https://www.ksda-bali.go.id/ca-batukahu/ diakses pada tanggal 4 Juni 2021.
Boyce PC, Sookchaloem D, Hetterscheid WLA, Gusman G, Jacobsen N, Idei T, Du NV.
2012. Acoraceae and Araceae, in Flora of Thailand 11(2):1-221. The Forest Herbarium, Department of National Parks, Wildlife and Plant Conservation: Bangkok.
Boyce PC, Wong SY. 2012. The Araceae of Malesia I: Introduction. Malay. Nat. J. 64(1): 33-67.
Boyce PC, Wong SY. 2015. Compendium Genera Aracearum Malesianum. Aroideana 38(1): 40-177.
Buggeln RG, Meeuse BJD, Klima JR. 1971. The Control of Blooming in Sauromatum guttatum (Araceae) by Darkness. Can. J. Bot. 49: 1025-1031. DOI: 10.1139/b71-143.
Dakwale SR, Bhatnagar S. 1985. Insect-trapping behaviour and diel periodicity in Sauromatum guttatum Schott (Araceae). Curr. Sci. 54(14): 699–702.
http://www.jstor.org/stable/24088626.
Diakses tanggal 7 Juni 2021.
Gibernau M. 2003. Pollinators and Visitors of Aroid Inflorescences. Aroideana 26: 66-83.
Gibernau M. 2011. Pollinators and Visitors of Aroid Inflorescences: an addendum.
Aroideana 34: 70-83.
Gibernau M, Barabe D. 2002. Pollination ecology of Philodendron squamiferum (Araceae). Can. J. Bot. 80: 316–320. DOI: 10.1139/b02-006
Grayum MH. 1990. Evolution and Phylogeny of the Araceae. Ann. Missouri Bot. Gard. 77(4): 627-697. DOI: 10.2307/2399668
Hadacek F, Weber M. 2008. Club-shaped Organs as Additional Osmophores within the Sauromatum Inflorescence: Odour Analysis, Ultrastructural Changes and Pollination
Aspects. Plant Biol. 4(3): 367-383.
DOI:10.1055/s-2002-32335
Heng L, Hetterscheid WLA. 2010. Sauromatum Schott in Schott & Endlicher, Melet. Bot. 17. 1832, in Fl. China 23: 36-39.
http://www.efloras.org/florataxon.aspx?flora _id=2&taxon_id=129334. Diakses tanggal 7 Juni 2021.
Hentrich H, Kaiser R, Gottsberger G. 2010. Floral Biology and rReproductive Isolation by Floral Scent in Three Sympatric Aroid Species in French Guiana. Plant Biol. 12: 587–596. DOI: 10.1111/j.1438-
8677.2009.00256.x
Mayo SJ, Bogner J, Boyce PC. 1997. The Genera of Araceae. The Trustees, Royal Botanic Gardens: Kew.
McDougal KM, Moore JE, Fanning ME. 2016. Biology of Weird Plant: The Voodoo Lily (Sauromatum guttatum). Proceedings of the 37th Conference of the Association for Biology Laboratory Education (ABLE), 37, Article 47 in Tested Studies for Laboratory Teaching (K. McMahon, Editor).
Meeuse BJD. 1975. Thermogenic Respiration in Aroids. Ann. Rev. Plant Physiol. 26: 117126. DOI:
10.1146/annurev.pp.26.060175.001001
Meeuse BJD. 1978. The Physiology of Some Sapromyophilous Flowers. In: A.J. Richards (ed.). The Pollination of Flowers by Insects. Linnean Society Symposium Series. No. 6. Academic Press: London, 97-104.
Meeuse BJD, Buggeln RG. 1969. Time, Space, Light, and Darkness in the Metabolic Flare-up of the Sauromatum Appendix. Acta Bot.
Neerl. 18(1): 159-172. DOI: 10.1111/j.1438-8677.1969.tb00581.x
Meeuse BJD, Hatch MH. 1960. Beetle Pollination in Dracunculus and Sauromatum (Araceae). Coleopteris Bull. 14(3): 70-74.
http://www.jstor.org/stable/3999116.
Diakses tanggal 7 Juni 2021.
Odyuo N, Roy DK, Dey S, Mao AA. 2015. Sauromatum horsfieldii (Araceae – Areae): an addition to the Flora of India. Telopea 18: 227-232. DOI: 10.7751/telopea8886.
Putri DMS. 2018. Koleksi Rhododendron Sebagai Salah Satu Taman Tematik di Kebun Raya “Eka Karya” Bali. Buletin Udayana Mengabdi. 17(1): 1-6.
Seymour RS. 2001. Biophysics & Physiology of Temperature Regulation in Thermogenic Flowers. Biosci. Rep. 21(2): 223-236. DOI: 10.1023/a:1013608627084
Seymour RS, Schultze-Motel P. 1997. Heatproducing Flowers. Endeav. 21(3): 125-129. DOI: 10.1016/S0160-9327(97)80222-0
Siregar M, Undaharta NKE. 2014. Vegetasi Alami dan Perubahannya setelah 22 tahun (1986-2008) di Hutan Tanaman Altingia excelsa Noronha Candikuning-Bali. Berita Biologi 12(2): 191-202. DOI:
10.14203/beritabiologi.v13i2.693.
Siregar M, Undaharta NKE. 2018. Tree Standing Dyamnics after 30 Years in A Secondary Forest of Bali, Indonesia. Biodiversitas 19(1): 22-30.
Skubatz H, Kunkel DD, Howald WN, Trenkle R, Mookherjee B. 1996. The Sauromatum guttatum Appendix as an Osmophore: Excretory Pathways, Composition of Volatiles and Attractiveness to Insects. New Phytol. 134: 631-640. DOI: 10.1111/j.1469-8137.1996.tb04928.x
Skubatz H, Nelson TA, Meeuse BJD, Bendich AJ. 1991. Heat Production in the Voodoo Lily (Sauromatum guttatum) as Monitored by Infrared Thermography. Plant Physiol. 95: 1084-1088. DOI: 10.1104/pp.95.4.1084
Yuzammi, Astuti IP, Suratman M. 2017. Sauromatum horsfieldii Miq. (Araceae): Koleksi Baru Kebun Raya Bogor. Warta Kebun Raya 15(1): 13-17.
10
Discussion and feedback