CA2+ INTRASELULER TERLIBAT DALAM MEKANISME PEMBUKAAN STOMATA AKIBAT PENGARUH AUXIN
on
JURNALBIOLOGI XII (1):19-22
ISSN: 14105292
CA2+ INTRASELULER TERLIBAT DALAM MEKANISME PEMBUKAAN STOMATA AKIBAT PENGARUH AUXIN
made pharmawati, made ria defiani. Ni luh arpiwi
Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Udayana, Kampus Bukit Jimbaran. Email:
pharmawati@hotmail. com
INTISARI
Penelitian ini bertujuan untuk mengetahui peranan Ca2+ intraseluler dalam mekanisme gerakan stomata yang disebabkan oleh auxin. Auxin dalam bentuk LAA dan NAA menyebabkan peningkatan pembukaan stomata. Penambahan EGTA, Ruthenium red dan Procaine yang merupakan modulator Ca2+ menghambat induksi pembukaan stomata oleh hormon tumbuhan. EGTA dan Procaine menghambat pembukaan stomata yang diinduksi oleh LAA dan NAA pada konsentrasi 100 uM dan 1 mM. Ruthenium red memberikan pengaruh pada konsentrasi yang lebih rendah yaitu 10 uM, 50 uMand 100 uM pada induksi pembukaan stomata oleh LAA, sedangkan pada induksi pembukaan stomata oleh NAA, Ruthenium red berpengaruh pada konsentrasi 10 |uM, 50 uM, 100 |uM dan 1 mM). Hal ini menunjukkan bahwa dalam kerjanya, IAA dan NAA tergantung pada Ca2+
Kata kunci: kalsium, auxin, stomata
ABSTRACT
1
Research was done to elucidate the role of Ca2+ intracellular in stomatal movement induced by auxin. Two types of auxin (IAA and NAA) increased stomatal opening and the increase was concentration dependent. The addition of EGTA, Ruthenium red and Procaine as the modulator of Ca2+ concentration , inhibited stomatal opening induced by auxin. EGTA and procaine significantly inhibited somatal opening at 100 uM and 1 mM. Ruthenium red worked at lower concentrations which were 10 uM, 50 uM and 100 uM on LAA-induced stomatal opening, while on NAA-induced stomatal opening, Ruthenium red had an effect at 10 uM, 50 uM, 100 uM dan 1 mM. It is suggested that Ca2+ involves in stomatal opening induced by LAA and NAA as signaling agent.
Keywords: calcium, auxin, stomata
PENDAHULUAN
Stomata merupakan bagian pada epidermis organ tumbuhan yang terdiri dari suatu celah yang dikelilingi oleh sel khusus yang disebut sel penjaga. Stomata sangat penting bagi tumbuhan karena pori stomata merupakan tempat terjadinya pertukaran gas dan air antara atmosfer dengan system ruang antar sel yang berada pada jaringan mesofil di bawah epidermis. Hal ini menyebabkan stomata sangat berperan dalam proses transpirasi dan fotosintesis (Moore et al, 1998).
Pergerakan pori stomata disebabkan oleh perubahan pada volume sel penjaga yang diatur oleh keluar masuknya ion K+ dan ion-ion lain dari dan ke sel penjaga selama proses pembukaan dan penutupan stomata. Sehubungan dengan fungsi stomata untuk menjaga hidrasi dan pertukaran gas, maka perubahan volume sel penjaga merupaka respon terhadap factor-
faktor lingkungan dengan melalui sinyal-sinyal yang kompleks (Moore etal., 1998, Hopkins, 1999).
Cahaya, konsentrasi CO2, kelembaban dan hormone tumbuhan merupakan beberapa factor yang mempengaruhi membuka dan menutupnya stomata (Kearn dan Assmann, 1993, Hopkins, 1999). Cahaya menyebabkan membukanya stomata sedangkan keadaan gelap, peningkatan konsentrasi CO2 dan turunnya kelembaban menyebabkan menutupnya stomata. Di antara sekian banyak hormone tumbuhan, ABA (abscisic acid) dan auxin merupakan hormone tumbuhan yang terkenal mempunyai pengaruh pada pergerakan stomata. ABA menyebabkan menutupnya stomata sedangkan auxin menyebabkan membukanya stomata (Hopkins, 1999).
Dalam proses fisiologi seluler terdapat beberapa sinyal yang berfungsi sebagai perantara yang dikenal dengan istilah second messenger yang terjadi setelah terbentuk kompleks hormone-reseptor (Assmann,
Naskah ini diteritna 1 Februari 2008, disetujui Mei 2008.
1993). Pada hewan, keberadaan second messenger dan aturan mainnya dalam sel telah banyak diketahui. Pada tumbuhan proses biokimia yang mengikuti kompleks hormone-reseptor belum banyak diteliti. Walupun dilaporkan bahwa pH intraseluler (Gehring et al, 1997), siklik nukleotida (Pharmawati et al, 1999), tetapibelum jelas apakah semua hormone tumbuhan menggunakan Ca2+, pH intraseluler dan siklik nukleotida sebagai perantara untuk menghasilkan respon fisiologis. Oleh karena itu dilakukan penelitian yang bertujuan untuk mengetahui apakah IAA (indole-3-acetic acid) dan NAA (naphtalen-3-acecic acid) yang merupakan kelompok hormone auxin dalam menginduksi pembukaan stomata pada epidermis bawah daun Vicia faba menggunakan Ca2+ sebagai bagian dari rangkaian sinyal kompleks. Serta untuk menguji apakah Ca2+ dikeluarkan dari tempat penyimpanan seluler (vakuola dan retikulum endoplasma) sebagai respon terhadap auxin.
MATERI DAN METODE
Bahan tanaman
Biji Vicia faba varietas early long pod direndam dalam air selama 12 jam dan dikecambahkan pada kertas sating basah dalam cawan petri. Setelah kecambah tumbuh, selanjutnya kecambah ditanam dalam pot dengan media tumbuh tanah dan disiram 1 kali sehari.
Bioassay untuk respon stomata terhadap auxin
Lapisan epidermis bawah daun Viciafaba yang berumur 4 minggu dikupas dengan penggunakan forcep. Daun diambil pada menjelang pagi hari untuk mengurangi adanya stomata yang sudah terbuka akibat pengaruh cahaya. Lapisan epidermis bawah dipotong dengan ukuran 0.5 cm x 0.5 cm dan diletakkan pada medium inkubasi (10 mM MES (pH 6.3), 50 mM KC1, 1 mM MgCk) dengan lapisan epidermis atas berada pada bagian atas. Auxin yang digunakan adalah IAA dan NAA dengan kosentrasi akhir 1 uM, 10 uM, 50 uM, 100 uM dan 1 mM ditambahkan ke dalam medium inkubasi. Inkubasi dilakukan selama 60 menit dalam keadaan terang.
Pemberian pengkelat dan penghambat saluran Ca2+
Ke dalam medium inkubasi ditambahkan EGTA (ethyleneglycol-bis (3-aminoethyl ether)-N,N,N',N' tetraacetc acid) dengan konsentrasi 1 u,M, 10 uM, 100 u,M dan 1 mM. Ruthenium red dan procaine ditambahkan secara terpisah dengan konsentrasi akhir 10 uM, 50 uM, 100 uM dan 1 mM.
Pengukuran pori stomata
Untuk tiap perlakuan dilakukan 3 kali ulangan, dan tiap ulangan terdiri dari 3 potongan epidermis bawah daun Viciafaba. Pengamatan pengukuran stomata dilakukan secara blind test (tidak diketahui sampel mana
yang mendapat perlakuan tertentu untuk menghindari bias dalam pengukuran). Setelah selesai tahap inkubasi, potongan epidermis diletakkan pada gelas benda dan diamati dibawah mikroskop. Diameter pori stomata diukur dengan menggunakan micrometer yang sudah dikalibrasi. Pada masing-masing ulangan 30 stomata diukur dari tiap potongan epidermis dari 3 lokasi bidang pengamatan yang berbeda. Data dianalisis dengan menghitung standar eror (SE) dan membandingkan antar perlakuan (http://www.ncsu.edu/labwrite/res/gt/ gt-stat-home.html)
HASIL DAN PEMBAHASAN
Hasil
Pembukaan stomata oleh IAA dan NAA
Pada Gambar 1 terlihat bahwa auxin mempengaruhi gerakan stomata. Konsentrasi LAA 1 uM menyebabkan membesarnya pembukaan stomata, tetapi tidak berbeda dengan kontrol. Pemberian IAA sebesar 10 uM menyebabkan peningkatan pembukaan stomata yang berbeda dengan kontrol. Pembukaan stomata terus meningkat sampai pemberian IAA 1 mM tetapi tidak berbeda dengan pemberian konsentrasi 50 u,M dan 100 uM.
Pada perlakuan NAA, pemberian NAA dengan konsentrasi 50 uM menghasilkan pembukaan stomata yang maksimum (Gambar 1). Pada pemberian NAA 1 mM, pembukaan stomata mendekati tingkat yang sarna dengan kontrol.
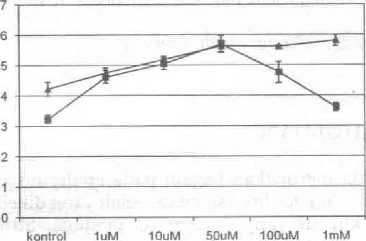
Gambar 1. Pembukaan stomata yang diinduksi olah IAA dan NAA. Setiap data point merupakan rata-rata ±SE dihitung dari 3 percobaan terpisah.
Pengaruh pengkelat dan penghambat saluran Ca2+
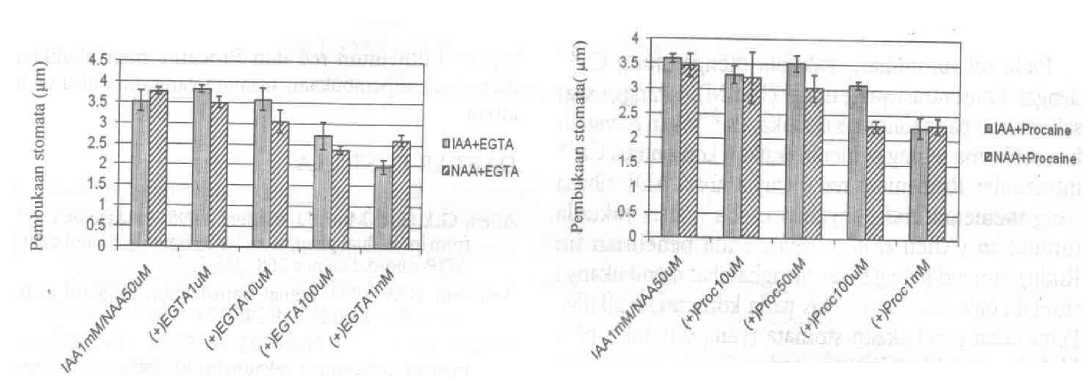
Respon pembukaan stomata yang diinduksi oleh IAA dan NAA terhadap EGTA sebagai Ca2+ kelator dapat dilihat pada Gambar 2. Pembukaan stomata yang diinduksi oleh IAA mengecil secara signifikan pada konsentrasi EGTA 100 uM. Konsentrasi EGTA 1 mM memberikan penghambatan terbesar. Pada pembukaan stomata yang diinduksi oleh NAA, EGTA menghambat pembukaan stomata pada konsentrasi 100 u,M dan ImM.
Gambar 3 menunjukkan pengaruh Ruthenium red
Gambar2. Pengaruh EGTA terhadap kemampuan lAAdan NAA untuk membuka stomata Setiap data point merupakan rata-rata ±SE dihitung dari 3 percobaan terpisah
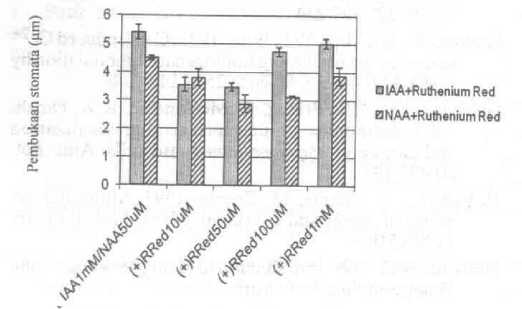
Gambar 3. Pengaruh Ruthenium red terhadap pembukaan stomata yang diinduksi oleh IAA dan NAA. Setiap data point merupakan rata-rata ±SE dihitung dari 3 percobaan terpisah.
Ca2+ Intraseluler Teiiibat dalam Mekanisme Pembukaan Stomata Akibat Pengaruh Auxin [Made Pharmawati, Made Ria Defiani, Ni Luh Arpiwi]
DIAA+EGTA
UNAAtEGTA
OIAAtProcaine
BNAAtProcaIne
terhadap induksi pembukaan stomata oleh IAA dan NAA. Ruthenium red pada konsentrasi 10 uM dan 50 uM menghambat pembukaan stomata yang diakibatkan oleh IAA, sedangkan pada pembukaan stomata yang diinduksi oleh NAA, Ruthenium red mengecilkan pembukaan stomata pada konsentrasi 10 u, 50 uM, 100 u.M dan 1 mM.
Pengaruh pemberian Procaine dapat dilihat pada Gambar 4. Pada induksi pembukaan stomata oleh IAA, pemberian Procaine pada konsentrasi 100 u.M dan 1 mM menyebabkan mengecilnya pembukaan stomata. Pada induksi pembukaan stomata oleh NAA, konsentrasi Procaine 100 uM dan 1 mM menyebabkan mengecilnya pembukaan stomata.
Pembahasan
Hormon tumbuhan berperan sebagai pengatur kesetimbangan air melalui pengaruhnya terhadap gerakan stomata. Dari penelitian ini terlihat bahwa auxin baik IAA maupun auxin sintetis NAA, menginduksi pembukaan stomata, dan pembukaan stomata tersebut tergantung pada konsentrasi yang digunakan. Efek yang dihasilkan oleh auxin berasal dari teraktifkannya bermacam-macam gen dan dari perubahan transport
Gambar4. Pengaruh Procaine pada pembukaan stomata yang diinduksi oleh IAA dan NAA. Setiap data point merupakan rata-rata ±SE dihitung dari 3 percobaan terpisah.
ion (Hobbie, et al., 1994). Perubahan transport ion terjadi karena perubahan aktivitas plama membrane H+-ATPase atau perubahan aktivitas saluran ion (Claussen, et al, 1997).
IAA sampai konsentrasi \ mM tetap meningkatkan pembukaan stomata. Respon stomata terhadap konsentrasi IAA terlihat mendekati linier (Gambar 1). Menurut Levitt et al., (1987), IAA pada konsentrasi 10 uM sampai 1 mM menstimulasi pembukaan stomata.
Respon stomata terhadap NAA memiliki puncak pada konsentrasi 50 uM. Konsentrasi NAA diatas 50 uM mengakibatkan berkurangnya pembukaan stomata. Menurut Assmann (1993) auxin bisa mengakibatkan pembukaan atau penutupan stomata. Penutupan stomata oleh auxin berasosiasi dengan pengaturan auxin terhadap tipe R saluran anion (R=rapid). Saluran anion tipe R ini teraktivasi dengan cepat sebagai respon terhadap depolarisasi dan terdeaktivasi dengan cepat sebagai respon terhadap hiperpolarisasi.
Perubahan konsentrasi Ca2+ dalam sitosol (Ca2+ intraseluler) telah diketahui berpengaruh dalam banyak jalur signaling, antara lain pada signaling pertumbuhan tabung polen, pertumbuhan ujung akar, respon terhadap stress lingkungan maupun pada penutupan stomata oleh ABA (Absisic acid) (Chalivendra et al., 1994). Konsentrasi Ca2+ intraseluler juga ditemukan meningkat akibat luka (Saniewski et al., 2006). Pada penelitian ini dibuktikan bahwa perubahan konsentrasi Ca2+ intraseluler juga berpengaruh pada pembukaan stomata yang diinduksi oleh auxin. Zat-zat yang mengubah konsentrasi Ca2+ intraseluler yaitu EGTA, Ruthenium red dan Procaine menyebabkan menurunnya pembukaan stomata yang diinduksi oleh auxin.
Gerakan stomata dipengaruhi oleh perubahan konsentrasi Ca2+ internal. Konsentrasi Ca2+ intraseluler meningkat mendahului respon stomata terhadap hormone (Irving et al., 1992). Hal ini disebabkan oleh dikeluarkannya Ca2+ dari tempat penyimpanan seluler. Ruthenium red dan Procaine merupakan zat yang menghambat pengeluaran Ca2+ dari penyimpanan
intraseluler. Pada sel hewan Ruthenium red dan Procaine menghambat reseptor ryanodine yang berhubungan dengan induksi pengeluaran Ca2+ (Galione et al., 1991).
Pada sel tumbuhan, vakuola mengandung Ca2+ dengan konsentrasi yang tinggi (lOmM), sehingga saat saluran ion pada tonoplas terbuka, Ca2+ akan mengalir ke sitoplasma sehingga meningkatkan konsentrasi Ca2+ intraseluler. Ruthenium red menghambat cADP-ribosa yang memerantarai pengeluaran Ca2+ dari vakuola tumbuhan (Allen et al., 1995). Pada penelitian ini Ruthenium red paling besar menghambat membukanya stomata oleh IAA dan NAA pada konsentrasi 50 uM. Penurunan pembukaan stomata (yang diinduksi oleh IAA dan NAA) akibat pemberian 50 uM Ruthenium red berturut-turut sebesar 1.93 um dan 1.61 um. menurut Pineros dan Tester (1997), penghambatan oleh Ruthenium red terjadi pada konsentrasi rendah yaitu 10 uM, 15 uM sampai 50 uM. tetapi Pineros dan Tester (1997) juga menjelaskan bahwa sensitivitas saluran Ca2+ pada membrane tumbuhan terhadap zat kimia bervariasi pada tumbuhan yang berbeda.
Procaine bekerja menghambat saluran yang melepaskan Ca2+ dari retikulum sarkoplasma dengan cara memperpanjang waktu menutupnya saluran ion (Zahradnikova dan Palade, 1993). Dengan tertutupnya saluran ion, maka Ca2+ tidak dapat dikeluarkan dari reticulum sarkoplasma ke sitoplasma, sehingga konsentrasi Ca2+ intraseluler menjadi rendah, dan hal ini menghambat pembukaan stomata oleh auxin.
EGTA merupakan zat yang mengkelat Ca2+, sehingga menurunkan konsentrasi Ca2+. Pada penelitian ini, EGTA pada 100 uM dan 1 mM menyebabkan menurunnya pembukaan stomata akibat induksi IAA dan NAA. Pengaruh EGTA juga dilaporkan oleh Webb et al. (1996) yang menemukan bahwa EGTA menurunkan konsentrasi Ca2+ intraseluler dan menghambat induksi penutupan stomata oleh COz.
Dari penelitian ini terlihat Ca2+ terlibat dalam gerakan stomata akibat pemberian hormon. Hal ini dibuktikan dari menurunnya pembukaan stomata oleh hormon jika diberikan zat-zat yang mengkelat Ca2+ atau zat-zat yang memblok saluran ion. Berarti auxin bekerja membuka stomata melalui mekanisme meningkatnya konsentrasi Ca2+. Dengan dihambatnya pengeluaran Ca2+ dari penyimpanan seluler maka induksi pembukaan stomata oleh auxin menjadi terhambat.
KESIMPULAN
Dari penelitian ini dapat disimpulkan bahwa auxin mempengaruhi gerakan stomata dengan menginduksi pembukaan stomata. Pengaruhnya ditentukan oleh konsentrasi yang digunakan, dimana IAA membuka stomata Viciafaba pada konsentrasi 10 uM sampai 1 mM, sedangkan NAA memberikan respon maksimum pembukaan stomata pada konsentrasi 50 uM.
Ca2+ terlibat dalam mekanisme gerakan membuka stomata yang diinduksi oleh auxin. Pengkelatan Ca2+ dengan EGTA dan penggunaan inhibitor saluran Ca2+ seperti Ruthenium red dan Procaine menyebabkan
menurunnya pembukaan stomata yang diinduksi oleh auxin.
DAFTAR PUSTAKA
Allen, G.J., S.R. Muir, D. sanders. 1995. Release of Ca2+ from individual plant vacuoles by both InsP3 and cyclic ADP-ribose. Science 268: 735-737
Assmann, S.M. 1993. Signal transduction in guard cells. Annu. Rev. Cell Biol. 9:345-375
Chalivendra, C. S., J. Zhang, M. M. Sachs. 1994. Involvement of intracellular calcium in anaerobic gene expression and survival of maize seedlings. Plant Physiol. 105: 369-376
Claussen, M., H., Luthen, M. Blatt, M. Bottger. 1997. Auxin-induced growth and its linkage to potassium channels. Planta 201:227-234
Galione, A., H.C. Lee, W.B. Busa. 1991. Ca2+ induced Ca2+ release in sea urchin egg homogenates: modulation by cyclic ADP-ribose. Science 253:1143-1146
Gehring, C.A., H.R. Irving, R. McConchie, R.W. Parish. 1997. Jasmonate induce intraselullar alkalization and closure of Paphiopedilum guard cells. Ann. Bot. 80:485-489
Hobbie, L., C. Timpte, M. Estelle. 1994. Molecular genetics of auxin and cytokinin. Plant. Mol. Biol. 26: 1499-1519
Hopkins, W.G. 1999. Introduction to plant physiology. John Wiley and Son, New York.
Irving, H.R., C.A. Gehring, R.W. Parish. 1992. Changes in cytosolic pH and calcium of guard cells precede stomatal movements. Proc. Natl. Acad. Sci. USA 88: 1790-1794
Kearns, E.V., S.M. Assmann. 1993. The guard cells environment connection.. Plant Physiol. 102:711-715
Levitt, L.K., D.B. Stein, B. Rubinstein. !987. Promotion of stomatal opening by indoleacetic acid and ethrel in epidermal strips of Viciafaba L. Plant Physiol. 85: 318-321
Moore, R., W.D. Clark, D.S. Vodopich. 1998. Botany. McGraw-Hill Companies Inc., USA.
Pharmawati, M., T. Billington, C.A. Gehring. 1999. Sto-matal guard cell responses to kinetin and natriuretic peptides are cGMP-dependent. Cell. Mol. Life Sci. 54:272-276
Pineros, M., M. Tester. 1997. Calcium channels in higher plant cells: selectivity, regulation and pharmacology. J. Exp. Bot. 48: 551-577
Saniewski, M., M. Horbowicz, A. Saniewska. 2006. The effect of Ruthenium red, a Ca++ channel blocker, on a red pigment formation in mechanically wounded scales of Hippeastrum x hybr. hort., and on the growth and development of Phoma narcissi. J. Fruit Ornam. Plant Res. 14:211-222
Webb, A.A.R., M.R. McAinsh, T.A. Mansfield, A.M. Hetherington. 1996. Carbondioxide induces increases in guard cell cytosolic free calcium. Plant J. 9: 297-304
Zahradnikova, A., P. Palade. 1993. Procaine effects on single sarcoplasmic reticulum Ca2+ release channels. Biophys. J. Biophysical Society 64:991-1003
Discussion and feedback