Virulensi Virus Newcastle Disease Isolat Lapang Berdasarkan Analisis Bioinformatika Gen Protein Hemaglutinin - Neuraminidase
on
Jurnal Ilmu dan Kesehatan Hewan, Februari 2015
Vol 3 No 1: 17-28
Virulensi Virus Newcastle Disease Isolat Lapang Berdasarkan Analisis Bioinformatika Gen Protein Hemaglutinin - Neuraminidase
Virulence of Field Isolates of Newcastle Disease Virus Based on Bioinformatic Analysis of Hemagglutinin-Neuraminidase Protein Genes
Fedry Rell1, Anak Agung Mirah Adi2, I Gusti Ngurah Kade Mahardika1,3* 1Laboratorium Biomedik dan Biologi Molekuler FKH-Unud, Denpasar 2Laboratorium Patologi Veteriner FKH-Unud, Denpasar 3Direktur Indonesian Biodiversity Research Center Unud, Denpasar *Corresponding author: gnmahardika@gmail.com
ABSTRACT
Newcastle disease virus (NDV) is very contagious disease agent, and causes many outbreaks trough out the country of Indonesia. This study was conducted to determine the virulence of Bali field isolate of NDV based on haemaglutinin-neuraminidase (HN) protein genes. Four field isolates from different locations were propagated at fertilized chicken eggs of 9 days old. Allantoic fluid was harvested and NDV was confirmed using standard haemaglutination (HA) and haemaglutination inhibition test (HI) methods. The fragment of HN protein gene was amplified using RT-PCR. The product was sequenced using Big-Dye termination method. All four isolates grew well with the titer of 25-29 HA unit and could be confirmed using HI test. The HN genes, however, exhibited variations at its 3-D structure and hydrophobicity between the virus that previously circulated in Indonesia and vaccine virus. It is concluded that all four Bali’s isolates under this study are virulent VND of the genotype VII. Further testing is needed to justify the best formula of NDV vaccine to be used trough out Indonesia.
Key words : Newcastle disease virus, genotype VII, HA, HI, RT-PCR, vaccine
ABSTRAK
Virus Newcastle disease (VND) merupakan agen penyakit yang sangat kontagius dan setiap tahun mewabah pada beberapa peternakan unggas di Indonesia. Tujuan penelitian ini adalah untuk menentukan virulensi VND isolat lapangan secara molekuler berdasarkan gen penyandi protein hemaglutinin-neuraminidase. Empat isolat VND yang berasal dari lokasi berbeda diperbanyak pada telur ayam bertunas (TAB) umur 9 hari melalui ruang allantois. Cairan alantois dipanen dan dilanjutkan dengan uji hemaglutinasi (HA) baku. Konfirmasi dilakukan dengan uji hambatan hemaglutinasi (HI) baku. Fragmen gen protein HN diamplifikasi dengan RT-PCR. Hasil RT-PCR divisualisasikan pada gel agarosa 1% dan disekuensing dengan Big-Dye termination method. Hasil uji HA keempat isolat VND memiliki titer kisaran 25-29 dan dapat dikonfirmasi dalam uji HI.Analisis sekuen asam amino protein HN menunjukkan variasi dalam struktur tiga dimensi dan hidrofobisitas antar VND yang pernah bersirkulasi di Indonesia dan virus vaksin. Dari hasil penelitian dapat disimpulkan bahwa 4 isolat VND adalah virulen dengan genotipe VII. Uji lanjutan perlu dilakukan untuk mendapatkan formulasi vaksin terbaik yang dapat digunakan di seluruh Indonesia.
Kata kunci : virus Newcastle disease, genotipe VII, HA, HI, RT-PCR, vaksin
PENDAHULUAN
Newcastle disease (ND) merupakan penyakit virus yang sangat kontagius yang menyebabkan penurunan produksi dan kerugian ekonomi pada peternakan ayam (Alexander, 2000; Dortmans, 2011).Penyakit
ND bersifat endemik di Indonesia dan ditemukan di berbagai daerah (Saepulloh dan Darminto, 2005). Sejauh ini penanggulangan wabah ND dilakukan dengan program vaksinasi namun sampai sekarang wabah ND tetap menjadi masalah yang serius di industri peternakan ayam (ADHPI, 2011; Xiao et al.,
2012). Sesuai dampak yang ditimbulkan maka Newcastle disease merupakan salah satu penyakit unggas yang termasuk daftar A oleh Office International des Epizooties (OIE) (Alaxander, 2000).
Penyakit ND disebabkan oleh virus Newcastle disease (VND) yang disebut Avian paramyxovirus serotype 1 (APMV-1), genus Avulavirus, famili Paramyxoviridae (OIE, 2012). Genom virus ND tersusun atas RNA berserat tunggal, polaritas negatif dengan panjang ± 15.200 nukleotida (nt). Genom itu menyandi 6 protein struktural, yaitu nukleoprotein(NP), phosphoprotein (P), matrix protein (M), fusion protein (F), hemaglutinin-neuraminidase protein (HN) dan large polymerase (L) (Alexander, 1987; 2000). Selain protein struktural diatas, terdapat dua protein non struktural yang disandi oleh gen P, yaitu protein V dan M (Yusoff dan Tan, 2001).
Avian paramyxovirus serotype1 menginfeksi sebagian besar spesies unggas, namun ayam merupakan spesies yang paling peka. Berdasarkan virulensi VND dalam membunuh ayam dan telur ayam bertunas, virus ini dikelompokan menjadi lima subtipe yaitu : 1) tipe velogenik viscerotropik adalah tipe VND yang menyebabkan kematian akut dengan perdarahan pada saluran pencernaan dan banyak ditemukan di Asia; 2) tipe velogenik neurotropik yang menimbulkan gangguan pernapasan dan saraf dengan tingkat kematian yang tinggi pada kebanyakan unggas di Amerika; 3) tipe mesogenik yang menimbulkan gangguan pernapasan dan saraf dengan mortalitas rendah; 4) tipe lentogenik yang menimbulkan penyakit ringan dengan gangguan pernapasan; 5) tipe asimptomatik enterik yang bereplikasi pada saluran pencernaan namun tidak menimbulkan kematian (Alexander, 1987; 2000). Umumnya, virulensi VND ditentukan dengan uji patogenisitas menggunakan mean death time (MDT), intravenous pathogenicity index (IVPI), dan Intracerebral pathogenicity index (ICPI) (OIE, 2004; Dortmans et al., 2011). Seiring dengan perkembangan ilmu pengetahuan dan bioteknologi, virulensi VND dapat ditentukan secara molekuler
berdasarkan sekuen gen penyandi protein F dan HN menggunakan software yang tersedia (Ke et al., 2010; Zhang et al., 2011).
Infeksi APMV-1 sampai sekarang masih sulit diberantas. Vaksinasi merupakan upaya pencegahan paling utama dari infeksi virus ND. Vaksin virus ND dapat berasal dari tipe velogenik, mesogenik, dan lentogenik. Umumnya, vaksin yang banyak beredar di Indonesia dibuat dari tipe letogenik, yaitu virus La Sota dan Hitchner B1 asal Amerika, dan tipe asimptomatik enterik, yaitu virus ND Ulster dan PHY-LMV asal Eropa. Jauhnya jarak genetik dengan virus lokal akan berkorelasi pada tingkat protektifitas dari hasil vaksinasi dan menjadi sebab merebaknya wabah ND walaupun program vaksinasi lengkap dan berulang telah dilakukan (ADHPI, 2011).
Protein HN pada amplop VND berperan penting dalam proses infeksi sekaligus sebagai penentu virulensi serta merangsang pembentukan antibodi protektifitas pasca vaksinasi (Mahon et al., 2008). Protein HN terdiri atas HA yang berperan dalam perlekatan pertikel virus dengan reseptor pada sel target dan NA yang berperan dalam pelepasan virus dari sel terinfeksi. Selain itu, protein HN juga berperan sebagai promoter aktivasi protein F (McGinnes dan Morrison, 2006; Takimoto et al., 2000).
Gen penyandi protein HN tersusun ± 2031 nt yang memiliki open reading frame (ORF) yang menyandi beberapa asam amino. Sekuen asam amino protein HN terdiri dari domain – domain penting, seperti posisi pengikat reseptor (receptor binding), posisi glikosilasi (N-linked glycosylation), posisi antigenik (antigenic sites), transmembran domain, posisi C-terminal extension, dan epitop sel B (Ponnusamy et al., 2009). Posisi antigenik dan epitop sel B berperan untuk mengenali sistem imun hospes yang berkontribusi dalam menentukan keberhasilan pasca vaksinasi.Transmembran domain bersifat hidrofobik berperan dalam perlekatan protein HN. Posisi C-terminus extension berperan menentukan panjang asam amino protein HN (Yusoff dan Tan, 2001). Panjang asam amino protein HN dapat dipakai dalam
pengelompokan virus ND.Virus ND virulen memiliki panjang C-terminus extension pada posisi asam amino 571 (Oberdorfer et al., 2003). Homologi protein HN virus vaksin dengan virus lapang akan sangat berkontribusi terhadap keberhasilan vaksinasi. Penelitian ini ditujukan untuk menentukan virulensi virus ND isolat lapangan yang menimbuklan wabah di beberapa lokasi dan waktu di Bali dengan melihat variasi gen protein HN.
MATERI DAN METODE
Rancangan Penelitian
Jenis racangan penelitian yang dilakukan adalah penelitian deskriptif observasional. Sekuen asam amino protein HN VND isolat lapang dianalisis untuk mengetahui virulensi VND secara molekuler, struktur 3-dimensi, hidrofobisitas dan hubungan genetiknya dengan data sekunder yang di akses dari GenBank.
Lokasi dan Waktu Penelitian
Penelitian dilakukan di Laboratorium Biomedik dan Biologi Molekuler Fakultas Kedokteran Hewan, Universitas Udayana, pada bulan Desember 2013 sampai dengan Agustus 2014.
Objek Penelitian
Objek dalam penelitian ini adalah virus Newcastle disease isolat lapang dari beberapa tempat di pulau Bali pada tahun 2013 – 2014. Sebanyak empat VND yang diobservasi dalam penelitian ini yaitu VND/ayam/lokal/ Badung/Bali/2014, VND/ayam/ras/Klungk-ung/Bali/2014, VND/ayam/ras/Tabanan1/
Bali/2014, dan VND/ayam/ras/Tabanan2/ Bali/2014.
Teknik Pengambilan Sampel
Cara pengambilan sampel menggunakan teknik purposive sampling yaitu mengambil sampel berdasarkan kejadian wabah ND pada suatu tempat.
Variabel Penelitian
Variabel dalam penelitian ini adalah sekuen nukleotida gen penyandi protein HN.
Sekuen gen penyandi protein HN dianalisis untuk menentukan virulensi VND secara molekuler dan filogenetik dengan isolat dari data sekunder serta struktur 3-dimensi dan hidrofobisitasnya. Data lainnya adalah data sekunder berupa sekuen nukleotida gen penyandi fragmen protein HN pada posisi asam amino 6418-8133 dari Negara lain dan virus vaksin yang diakses dari GeneBank.
Prosedur Penelitian
Sampel virus Newcastle Disease
Sebanyak empat sampel VND isolat lapang yang diisiolasi dari kasus ND pada beberapa tempat di pulau Bali. Keempat VND isolat lapang yaitu VND/ayam/lokal/ Badung/Bali/2013 yang diisolasi dari kasus ND pada peternakan ayam lokal di Kabupaten Badung pada bulan Desember 2013, VND/ayam/ras/ Klungkung/ Bali/2014 yang diisolasi dari kasus ND pada peternakan ayam broiler di Kabupaten Klungkung pada bulan Januari 2014, VND/ayam/ras/Tabanan1/ Bali/2014 yang diisolasi dari kasus ND pada peternakan ayam broiler di Kabupaten Tabanan pada bulan Maret 2013, dan VND/ayam/ras/Tabanan2/Bali/2014 yang disolasi dari wabah ND pada peternakan ayam di Desa Payangan Kabupaten Tabanan pada bulan Mei 2014. Keempat sampel tersebut disimpan di Laboratorium Biomedik dan Biologi Molekuler Fakultas Kedokteran Hewan, Universitas Udayana.
Propagasi Virus Newcastle Disease
Keempat sampel VND isolat lapang dipropagasi pada pada telur ayam bertunas (TAB) umur 9-10 hari melalui ruang alantois (allantoic cavity). Propagasi pada TAB diawali dengan pengamatan telur (candling) untuk mengetahui keadaan embrio. Kemudian virus ND isolat lapangan diinjeksikan ke dalam ruang alantois sebanyak 0,1-0,2 ml/butir telur dengan menggunakan tuberculin syringe. Lubang bekas tusukan pada cangkang ditutup dan selanjutnya TAB diinkubasikan pada suhu 37°C.Pengamatan dilakukan setiap hari selama 4 hari.Selanjutnya cairan alantois
dipanen dari telur embrio yang mati dan dilanjutkan dengan uji serologi dan RT-PCR.
Uji Hemaglutinasi
Uji hemaglutinasi (HA) adalah salah satu uji serologi untuk mengatahui adanya virus yang menghemaglutinasi sel darah merah (SDM). Sampel yang positif ditandai dengan adanya aggregate SDM seperti pasir pada dasar sumuran mikroplat (Kencana et al., 2012). Uji HA dari keempat sampel VND isolat lapang mengacu pada prosedur standar dari OIE (2004) yang telah dimodifikasi oleh Kencana et al. (2012). Uji HA dilakukan dengan cara : menyiapkan mikroplat steril kemudian pada masing-masing lubang (1 – 12) ditambahkan dengan 25 µl NaCl 0,9%. Selanjutnya pada lubang pertama dan kedua ditambahkan antigen VND sebanyak 25 µl dan selanjutnya diencerkan berseri kelipatan dua mulai dari lubang kedua sampai lubang kesebelas dengan menggunakan pengencer mikro. Tambahkan NaCl 0,9% sebanyak 25 µl pada semua lubang (1 – 12). Kemudian ditambahkan ke dalam setiap lubang 50 µl suspensi sel darah merah ayam konsentrasi 1% kemudian diayak selama 30 detik. Selanjutnya dieramkan pada suhu kamar selama 1 jam dan reaksi diamati setiap 15 menit ada atau tidaknya reaksi hemaaglutinasi sel darah merah.
Uji Rapid Hambatan Hemaglutinasi
Uji rapid hambatan hemaglutinasi (HI) adalah uji cepat untuk mengetahui adanya hambatan hemaglutinasi virus oleh serum VND. Sampel yang positif ditandai dengan adanya gumpalan SDM pada dasar sumuran mikroplat. Rapid HI dilakukan dengan cara: menyiapkan mikroplat steril kemudian pada masing – masing lubang (1-12) ditambahkan 25 µl NaCl 0,9%. Lubang pertama sampai kesepuluh ditambahkan serum
VND.Selanjutnya ditambahkan antigen sebanyak 4 unit HA pada lubang pertama sampai kesebelas.Kemudian dieramkan pada suhu kamar salama 30 menit. Setelah itu, sebanyak 50 µl suspensi sel darah merah ayam kosentrasi 1% ditambakan pada semua lubang dan diayak kembali selama 30 detik.
Selanjutnya dieramkan pada suhu kamar selama 1 jam dan diamati setiap 15 menit ada tidaknya aglutinasi sel darah merah (Kencana et al., 2012).
Isolasi RNA Virus Newcastle Disease
Sebanyak 250 µl sampel dalam tabung eppendorf ditambahkan dengan 750 µl Trizol LS Reagent (Invitrogen). Campuran divorteks selama 1 menit kemudian diinkubasi pada suhu kamar selama 5 menit. Kemudian ditambahkan kloroform sebanyak 200 µl, lalu divorteks selama 15 detik dan diinkubasi pada suhu kamar selama 15 menit. Campuran selanjutnya disentrifuse dengan kecepatan 12.000 rcf selama 15 menit. Bagian aqueus dipindahkan ke dalam tabung eppendorf steril dan ditambahkan isopropil alkohol sebanyak 500 µl. Larutan diinkubasi pada suhu kamar selama 10 menit, kemudian disentrifuse dengan kecepatan 12.000 rcf selama 10 menit. Supernatan dibuang, lalu ditambahkan alkohol 70% sebanyak 1000 µl. Kemudian larutan disentrifuse dengan kecepatan 7.500 rcf selama 5 menit. Supernatan dibuang dan peletnya di air dry di inkubator selama 5-10 menit. Setelah peletnya agak mengering, lalu ditambahkan 20 µl treated water. Kemudian disimpan dalam freezer sampai digunakan.
Reverse Transcriptase – Polymerase Chain Reaction dan Elektroforesis
RNA VND yang telah diisolasi dengan menggunakan protokol Trizol dilanjutkan dengan teknik transkripsi balik dan diamplifikasi menggunakan reverse transcriptase – polymerase chain reaction (RT-PCR) standar dengan tiga pasang primer yang mengamplifikasi daerah protein HN pada posisi nukleotida 6418-8133.Informasi ketiga pasang primer dapat dilihat pada Tabel 1.
Teknik RT-PCR dilakukan dengan cara: tabung eppendorf diisi dengan R-mix (dNTP, MgSO4 dan buffer) sebanyak 5 µl, primer depan dan primer belakang sebanyak 0,6 µl, enzim SuperScripTM III onestep RT-PCR System with Platinum® Taq DNA Polymerase (Invitrogen) 0,25 µl, aquabides 2,55 µl dan RNA virus sebanyak 1 µl. Tabung eppendorf
kemudian dimasukkan dalam mesin thermocycler dan diprogram sebagai berikut: reverse RNA menjadi cDNA pada suhu 50°C selama 1 jam, pre-denaturasi pada suhu 95°C selama 7 menit dan denaturasi 94°C selama 45 detik. Selanjutnya proses annealing pada suhu 52°C selama 45 detik dan tahap extension pada suhu 72°C selama 1 menit. Tahap denaturasi, annealing, dan extension diulangi sampai 39 kali. Terakhir tahap penyempurnaan kerja enzim pada suhu 72°C selama 5 menit. Setelah tahapan penyempurnaan selesai mesin thermocycler kembali pada suhu 220C (Kencana et al., 2012). Produk RT-PCR disimpan ke dalam freezer sebelum dielektroforesis.
Tabd 1. Sgfajsi ⅛asa primer untuk untuk HieBgainplifikasi gen penyandi protein HN se besaran produk PCR yang diharapkan.
|
No Primer |
Sekuen basa |
Produk |
|
lh HN61F HN-OOB |
tgctaymtratgtay.∖arcaraaggc ctaaatt Gatggaacgc agagtag |
966⅛ |
|
2b NDVFF NDVFB |
Gttgcactcggataccctcat Acttcttgtaccaacgagggtc |
700⅛ |
|
3b HNtlOOF HN83B |
Ccatcttatctatcgaagtgtc Ggaccggagcccgccatgtcctacccgt |
8¾ |
Ker: cPeneliti
analisis jarak genetik menggunakan Pairwise Distance Method dan filogenetik menggunakan Phylogeny Construct/test Neighbor-Joining Tree Method dengan subtitusi nukleotida menggunakan Kimura 2-parameter model dari Mega5 (Tamura et al., 2011).
HASIL DAN PEMBHAASAN
Hasil
Isolasi dan Propagasi Virus Newcastle Disease
Uji HA dari keempat sampel VND menunjukkan reaksi positif (Gambar 1a) dengan titer kisaran 25 - 29 (Tabel 2). Uji Rapid HI menunjukkan reaksi positif bahwa keempat sampel merupakan virus ND karena tidak terjadi hemaglutinasi pada sumuran mikroplat yang berisi hiperimun serum ND (Gambar 1b).
Tabel 2. Hasil uji hemaglutinasi terhadap keempat sampel VND pada sel darah merah
1 VND/ayam/lokal/Badung/Bali/2013 NDl2
2 VND∕ayam⅛as∕KlungkungZBali∕2014 ND22
3 VND∕ayam∕ras∕Γabananl∕Bali∕2014 ND32
4 VND∕ayamΛas∕Γabanan2ZBali∕2014 ND42
Ket. Semua ernbτi□ TAB mati pada heri kedua pasca inokulasi.
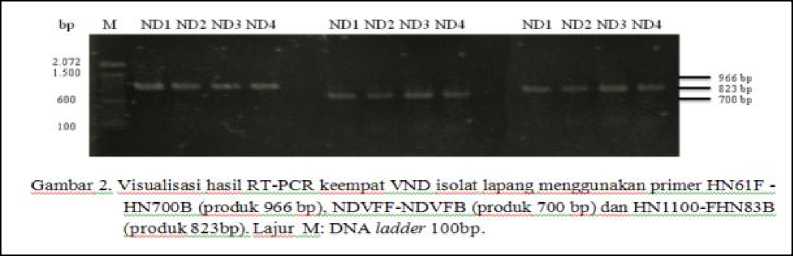
Hasil RT-PCR dari keempat VND isolat lapang kemudian dilanjutkan dengan elektroforesis dengan cara sebagai berikut: sebanyak 3 ul produk RT-PCR yang ditambahkan dengan blue juice (bromphenolblue dan cyline cyanol) sebanyak 1 µl yang dielektroforesis pada gel agarose 1% yang telah dicampur dengan etidium bromide pada mesin elektroforesis dengan tegangan 100 V (Volt) selama 30 menit. Hasil uji divisualisasikan dengan sinar ultraviolet dan didokumentasikan dengan kamera (Kencana et al., 2012).
Sekuensing dan Analisis Materi Genetik
Produk RT-PCR dikirim ke Berkeley Sequencing Fasility, University of California, USA untuk disekuensing. Runutan nukleotida hasil sekuensing VND isolat lapang dan turunan asam aminonya disepadankan dan dibandingkan dengan sekuen virus ND pada vaksin dan isolat dari Negara lain yang diakses dari GeneBank dengan Clustal W Method dari Mega5. Dilanjutkan dengan

Reverse Transcriptase – Polymerase Chain Reaction dan Elektroforesis
Setelah diidentifikasi dengan uji serologis dilanjutkan dengan isolasi RNA virus dan diuji dengan RT-PCR one step menggunakan tiga pasang primer yang menangkap gen penyandi protein HN. Identifikasi gen penyandi protein HN menggunakan pasangan primer HN61F dan HN700B untuk mengamplifikasi daerah
pertemuan dengan gen protein F, pasangan primer NDVFF dan NDVFB dipakai untuk mengamplifikasi daerah tengah gen protein HN, dan pasangan primer HN1100F dan HN83B dipakai untuk mengamplifikasi gen HN daerah pertemuan dengan gen penyandi protein L. Hasil penelitian menunjukkan bahwa keempat isolat positif VND (Gambar 2).
Sekuen Asam Amino Protein Hemaglutinin-Neuraminidase
Hasil analisis sekuen gen penyandi protein HN dari keempat VND isolat lapang menunjukkan daerah yang variabel terdapat pada posisi asam amino 1-80. Terdapat 13 residu cysteine berada pada asam amino 123, 172, 186, 196, 238, 247, 251, 344, 455, 461, 465, 531, dan 542. Tiga residu asam amino pada posisi 411 (E), 416 (R), dan 526 (Y) pada protein HN ditemukan pada sekuen asam amino protein HN VND isolat lapang. Residu glikosilasi (NxT/NxS) terdapat pada asam amino 119, 341, 433, 481, dan 538 kecuali pada isolat ND/ayam/ras/Tabanan2/Bali/ 2014. Kehilangan residu glikosilasi pada asam amino 119. Residu Arginine pada asam amino 174, 416, 498, dan 516 ditemukan pada
ke empat VND isolat lapang. Residu asam amino ujung C-terminal protein HN dari keempat VND isolat lapang yaitu 571.
Polimorfisme Sekuen Asam Amino Protein Hemaglutinin-Neuraminidase
Terdapat polimorfisme asam amino protein HN antar VND isolat lapang yaitu pada asam amino 59 (I/V), 119 (N/D), 288 (T/A), 309 (D/N) dan 569 (N/S). Jumlah situs polimorfisme asam amino protein HN antar VND yang pernah beredar di Bali terdapat sebanyak 48 situs (Gambar 3). Analisis pohon filogenetik berdasarkan sekuen nukleotida gen penyandi protein HN menunjukkan bahwa VND yang pernah beredar di Bali berbeda kluster dengan virus LaSota. Sama hal dengan VND yang pernah beredar di Bali terbagi menjadi dua kluster besar.Keempat VND isolat lapang (garis tebal) satu kluster dengan isolat Gianyar/013/10, Sragen/10, Banjarmasin/10, Kudus/10 dan isolat dari Pakistan/2011. Sedangkan NDV/Bali-1/07 dan Bali/020/10 juga terdapat satu kluster yang terpisah dengan keempat VND isolat lapang (Gambar4).

Hasil analisis jarak genetik berdasarkan sekuen asam amino protein HN menunjukkan jarak genetik VND yang pernah beredar di Bali dengan virus vaksin cukup jauh (Tabel 3) dengan kisaran 18 – 22%. Sedangkan jarak genetiknya dengan virus NDV-La Sota berkisar 21-22% (kolom yang berwarna
hitam). Jarak genetik yang satu kluster dengan keempat VND isolat lapang berkisar 0 - 1% (kolom yang berwarna merah). Sedangkan jarak genetik keempat VND isolat lapang dengan virus ND lain di Indonesia yang berbeda kluster berkisar 8 – 9% (kolom berwarna kuning).
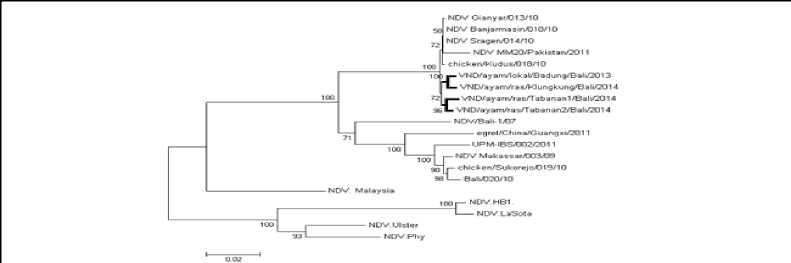
Garnbar 4. Pohon Eilogenik VNlD Berdasaifcan sekuen Iiukleotida gen penyandi protein HZN pada posisi 6418-8133 (1715 bp). Pohon Iilogenetik
dianalisis menggunakan PAy∕o⅛reray OorasfrwcJ/ JesJ A√⅛⅛AAor-7ozrazra,gr Tree AdrCtAoo! dari Mega5. Skala di bawali nieniii^ukfcan jarak antar sekuen. Angka pada pada pohon menunjukkan nilai Aootstrctp masing-masing c abang/r anting. PZeernp at VlNJD isolat _ lap an g ditandai dengan gaiis tebal.
Tabcl 3. Jarak genetik. VND yang pernah beredar di pulau Bali dan dari beberapa temp at. di Indone sia . dengan, .virus. vaksin.. yang di aks e s. dari. Ger? e B⅛r⅛.
men;
;a:
1 2 3
5 6 7 g
9 IO 11 12 13 14 15 16
Kgt Kolom yang berwarna Iiitani : jarak genetik dengan virus Lasota, kolom berwarna kuning : jarak genetik dengan virus ND di Indonesia yang berbeda kluster, kolom berwarna merah : jerak genetik yang satu kluster dengan keemρat isolat.
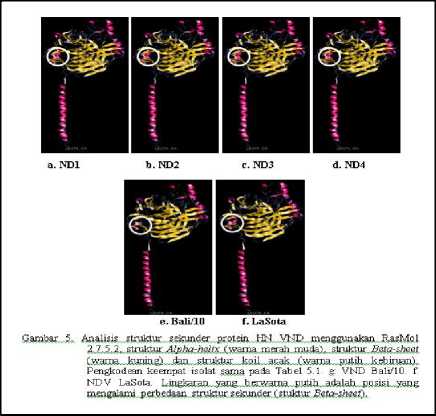
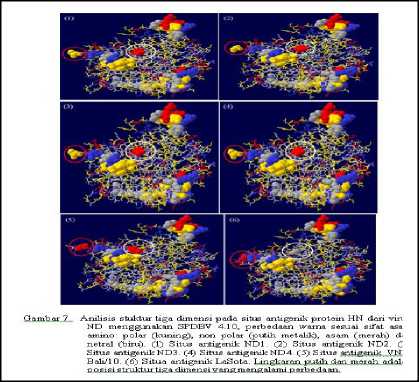
Analisis Struktur Tiga Dimensi
Hasil analisis stuktur sekunder protein HN keempat VND isolat lapang menunjukkan bahwa keempat isolat memiliki struktur yang sama. Struktur protein HN terbagi menjadi dua bagian utama yaitu bagian batang dan kepala yang berbentuk globular. Struktur protein HN ini terdiri dari: 1) struktur pilinan rantai asam-asam amino berbentuk seperti
spiral yang disebut struktur alpa-helix, 2) struktur lembaran-lembaran lebar yang tersusun dari sejumlah rantai asam amino yang disebut struktur Beta-sheet, dan 3) struktur seperti kawat yang disebut struktur koil acak (Gambar 5). Hasil perbandingan struktur sekunder dengan virus ND isolat Bali dan NDV LaSota ditemukan perbedaan struktur (lingkaran putih). Keempat isolat
VND memiliki struktur sekunder yang sama dengan LaSota, sedangkan dengan isolat Bali/10 memiliki perbedaan. Perbedaan tersebut terletak pada posisi asam amino 156158 yang membentuk struktur Beta sheet pada keempat isolat.
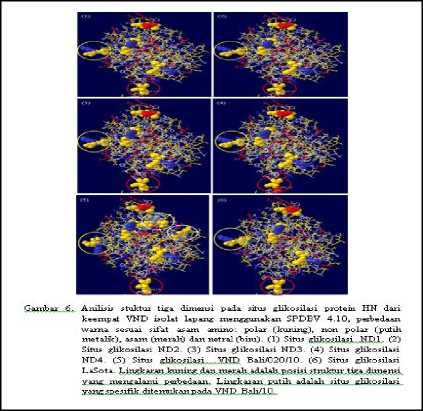
Hasil analisis struktur tiga dimensi pada situs glikosilasi protein HN antar keempat VND isolat lapang tidak menunjukkan perubahan struktur kecuali pada posisi asam amino 119 dariVND/ayam/ras/Tabanan2/Bali/ 2014 (ND4) yang merupakan situs polimorfisme (N/D) antar keempat VND isolat lapang yang menunjukkan perubahan struktur dan sifat pada asam amino tersebut dimana terdapat perbedaan warna kuning menjadi merah. Sedangkan hasil analisis struktur tiga dimensi antara keempat VND isolat lapang dengan isolat Bali/020/10 dan virus LaSota menunjukkan perbedaan struktur pada situs glikosilasi pada posisi 433-435 (lingkaran kuning).Selain itu, struktur tiga dimensi isolat Bali/020/10 mimiliki situs glikosilasi spesifik yang tidak ditemukan pada situs glikosilasi dari keempat VND isolat lapang dan LaSota (lingkaran putih) (Gambar 6).
Hasil analisis tiga dimensi situs antigenik protein HN antar keempat VND isolat lapang tidak menunjukkan perubahan struktur kecuali pada posisi asam amino 569 dari VND/ayam/ras/Klungkung/Bali/ 2014 (ND2) yang juga merupakan situs polimorfisme
(N/S) antar keempat VND isolat lapang yang menunjukkan perubahan struktur asam amino pada situs tersebut (Gambar 7). Sedangkan hasil analisis sruktur tiga dimensi situs antigenik protein HN antara keempat VND isolat lapang dengan isolat Bali/020/10 dan virus LaSota menunjukkan perbedaan struktur dan sifat dari asam aminonya.Situs antigenik yang mengalami perbedaan yaitu di posisi asam amino 494 (lingkran putih) dan 569 (lingkaran merah).
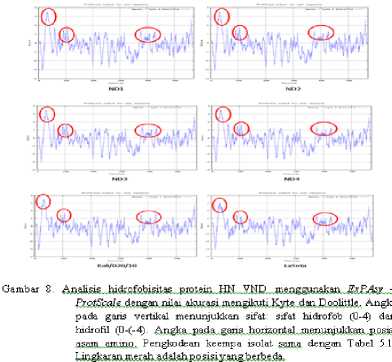
Analisis Hidrofobisitas
Hasil analisis hidrofobisitas protein HN antar keempat VND isolat lapang menunjukkan tidak ada perbedaan. Namun, hasil analisis hidrofobisitas antara keempat VND isolat lapang dengan isolat Bali/020/10 dan virus LaSota menunjukkan perbedaan hidrofobisitas pada beberapa asam amino
seperti di posisi 1–300 dan 400–500 (Gambar 8).
Pembahasan
Keempat isolat virus yang berhasil diisolasi dapat mengehemaglutinasi SDM. Berdasarkan kemampuan menghemaglutinasi SDM unggas pada uji HA, keempat isolat dapat dikelompokkan virus golongan Paramyxoviridae (misal ND ), Orthomyxoviridae (misal : AI), virus Infectious Bronchitis (IB) dan virus Egg Drop Syndrome'76 (EDS) (Miller dan Stanley, 1944; Kusmaedi, 2001). Pada uji HA titer dari keempat isolat berada dalam kisaran 25-29 dengan waktu inokulasi dan panen yang tidak berbedah jauh (Tabel 2 dan Gambar 1a).Perbedaan titer dari keempat isolat dapat disebabkan oleh maternal antibodi embrio pada masing-masing pada TAB (Yeo et al., 2003). Selain itu, virulensi virus ND mempengaruhi waktukematian embrio pada TAB dan titer virus (Khadzhiev, 1987; Absalὁn et al., 2012). Identifikasi virus dilanjutkan dengan uji Rapid HI menggunakan hiperimun serum ND dan AI.Hambatan hemaglutinasi SDM terjadi pada sumuran mikroplat yang berisi hiperimun ND sedangkan pada sumuran yang berisi hiperimun AI hambantan hemaglutinasi tidak terjadi (Gambar 1b).Hambatan hemaglutinasi terjadi karena reaksi pengikatan hiperimun serum ND terhadap antigenik virus ND yaitu protein HN (Alexander, 2000; McGinnes dan Morrison, 2003).Sehingga
protein HN tidak dapat berikatan dengan asam sialat yang bertindak sebagai reseptor SDM. Protein HN adalah protein dalam VND yang berperan untuk berikatan dengan asam sialat sebagai reseptor SDM yang menyebabkan hemaglutinasi atau pada sel lain dalam memulai proses infeksi (Couceiro et al., 1995; Panda, 2003).
Identifikasi virulensi dari keempat VND isolat lapang dilanjutkan dengan uji RT-PCR one step menggunakan pasangan primer FNDFP dan FNDBP dan produk RT-PCR disekuensing. Pasangan primer yang digunakan mengamplifikasi pada posisi nukleotida 4661-5016 yang merupakan daerah pemotongan (cleavage site) dengan produk 316 bp. Produk RT-PCR dielektroforesis pada gel agoresa 1% dan divisualisasikan dengan kamera digital menunjukkan hasil positif (hasil tidak dipublikasikan)
Gen penyandi protein HN VND yang multifungsional memiliki berat molekul 67.4 kDA (Yusoff et al., 1996). Berdasarkan analisis sekuen asam amino protein HN dari keempat VND isolat lapang terdapat 13 residu cysteine. Residu cysteine 123merupakan salah satu posisi asam amino yang variabel dengan virus La Sota (tryptophan/W) (data tidak dipublikasikan). Cysteine merupakan asam amino yang penting dalam interaksi ikatan disulfida intramolekul yang berperan dalam menstabilkan struktur protein HN VND (Takimoto et al., 2000). Ponnusamy et al. (2009) menyatakan bahwa residu cystine posisi asam amino 123 merupakan spesifik ditemukan pada VND virulen.Namun, pernyataan tersebut tidak sesuai dengan hasil analisis yang diperoleh karena virus vaksin NDV-PHY dan NDV-Ulster juga memiliki residu cysteine pada posisi 123.Kemungkinan residu cysteine 123 mempengaruhi struktur fungsional protein HN tapi tidak berdiri sendiri karena asam amino lain juga berkontribusi.
Hasil perbandingan residu asam amino ujung C-terminal protein HN VND terdapat lima jenis panjang residu asam amino (data tidak dipublikasikan). Variasi panjang protein HN tersebut dipengaruhi oleh lokasi kodon N-terminal protein HN yang berbeda-beda pada
setiap tipe VND sehingga panjang asam amino yang diekspresikan oleh gen HN bervariasi (Adi dan Astawa, 2014). Panjang residu asam amino keempat VND isolat lapang adalah 571.Laporan penelitian Oberdorfer et al. (2003) menyatakan bahwa panjang residu asam amino 571 spesifik ditemukan pada VND yang sangat virulen. Namun berdasarkan perbandingan panjang residu asam amino C-terminal protein HN, VND avirulen juga memiliki panjang residu asam amino 571 seperti NDV. Strain Mukteswar, NDV.Herts/33, NDV. Italien, dan lain-lain. Hal serupa sama dengan laporan penelitian Ke et al. (2010) yang menunjukkan bahwa VND avirulen memiliki panjang residu asam amino C-terminal 571. Namun dapat dipastikan panjang residu asam amino ujung C-terminal protein HN VND turut serta mempengaruhi virulensi virus karena semua VND virulen hanya memililki panjang protein HN yang sama yaitu 571.
Analisis situs polimorfisme asam amino protein HN menunjukkan bahwa keempat VND isolat lapang homogenus kecuali variasi dibeberapa situs (Gambar 3). Variasi antar keempat VND isolat lapang terletak pada posisi 59 (V/I), 119 (N/D) dan 569 (N/S). Kedua posisi terakhir merupakan posisi penting yang mempengaruhi peran protein HN dalam perlekatan dan pelepasan virus dari sel terifeksi.Posisi asam amino 119-121 yang memiliki residu asam amino asparagine (N) dan diakhiri serine/ threonine (S/T) merupakan daerah glikosilasi. Daerah glikosilasi pada protein HN berperan penting dalam afinitas terhadap reseptor pada sel target sehingga turut terlibat dalam mempengaruhi virulensi VND (Panda et al., 2004). Posisi asam amino 569 merupakan daerah antigenik protein HN yang menginduksi antibodi netralisasi dari host (Iorio dan Bratt, 1984; Iorio et al., 1991; Gong dan ZhiZhong, 2011).
Analisis pohon filogenetik juga menunjukkan bahwa VND yang pernah beredar di pulau Bali terpisah kluster dengan NDV (Gambar 4). La Sota. Selain itu, berdasarkan hasil analisis filogenetik kedua protein amplop diatas menunjukkan bahwa
pengelompokan kekerabatan VND selain menggunakan sekuen gen penyandi protein F juga dapat menggunakan sekuen gen penyandi protein HN.
Hasil analisis jarak genetik berdasarkan sekuen asam amino protein HN menunjukkan bahwa jarak genetik VND yang pernah beredar di pulau Bali dan dari beberapa tempat di Indonesia dengan virus vaksin cukup jauh (Gambar 5). Jarak genetik dengan virus vaksin kisaran 18 - 22%. Secara khusus, dengan virus vaksin LaSota sebagai vaksin komersial yang dipakai oleh sebagian besar para peternak di pulau Bali memiliki jarak genetik dengan VND lapang bali berada dalam kisaran 21 – 22 %. Perbedaan jarak genetik yang cukup jauh antara virus lapang dengan virus vaksin akan mempengaruhi keberhasilan vaksinasi. Keberhasilan
vaksinasi sangat ditentukan oleh pengenalan antibodi terhadap virus lapang pasca vaksinasi.Pengenalan antibodi dipengaruhi oleh homologi virus vaksin dengan virus lapang (Hughes, 2009; Perozo et al., 2012). Virus vaksin yang homolog dengan virus lapang akan merangsang produksi antibodi yang lebih cepat karena mengenali virus lapang yang menginfeksi. Pengenalan antibodi tersebut akan mempercepat netralisasi virus di dalam tubuh inang pada waktu infeksi awal. Sehingga, untuk membantu para peternak dalam pengendalian wabah ND diperlukan virus vaksin yang homologi dengan virus lapang.
Berdasarkan hasil analisis struktur tiga dimensi protein HN dari keempat VND isolat lapang tidak menunjukkan perbedaan kecuali pada analisis tiga dimensi daerah situs glikosilasi (119/D) dan antigenik (569/S). Pada ND/ayam/ras/Tabanan2/Bali/2014 yang mengalami variasi pada situs glikosilasi menunjukkan perbedaan tipe asam amino dan strukturnya (Gambar 6). Pada
ND/ayam/ras/Klungkung/Bali/2014 mengalami variasi pada situs antigenik menunjukkan perbedaan struktur asam amino pada posisi tersebut. Hasil analisis struktur tiga dimensi protein HN antara VND isolat lapang dengan isolat Bali/020/10 dan virus LaSota menunjukkan perbedaan struktur pada
situs yang sama kecuali pada dua posisi situs glikosilasi yang hanya ditemukan pada isolat Bali/020/10 dan situs antigenik pada posisi 494 (Gambar 7). Perbedaan ini memberikan gambaran bahwa apabila terjadi variasi asam amino akan mempengaruhi tipe dan struktur asam amino tersebut. Perbedaan tipe dan struktur asam amino akan mempengaruhi fungsi dari protein HN tersebut (Crennel et al., 2000; Takimoto et al., 2000).
Analisis hidrofobitas protein HN VND yang pernah beredar di pulau Bali dengan virus LaSota menunjukkan perbedaan pada beberapa posisi seperti di posisi antara 1 - 300 dan 400 – 500. Posisi asam amino 1 - 300 adalah daerah yang memiliki beberapa situs penting seperti bagian batang (1-143), situs transmembran (27-48), situs glikosilasi (119), dan epitop antigenik (193-201). Posisi asam amino 400 – 500 adalah daerah yang juga memiliki beberapa situs penting seperti situs glikosilasi (433, 481, dan 500) dan situs pengikat reseptor (401 dan 416). Kesemua posisi di atas mengalami peningkatan sifat hidrofob. Secara khusus pada situs transmembran mengalami peningkatan dari nilai hidrofob dibawah 3 menjadi ± 3,5. Laporan penelitian Botus et al. (2009) menyatakan bahwa Perbedaan hidrofobisitas protein HN mempengaruhi sifat/ fungsi antigenik dari virus ND. Perubahan sifat/ fungsi antigenik akan mempengaruhi virulensi VND. Analisis hidrofobisitas ini menunjukkan bahwa adanya variasi asam amino menyebabkan perubahan nilai hidrofobisitas.
SIMPULAN
Berdasarkan hasil penelitian disimpulkan bahwa keempat VND isolat lapang adalah VND virulen yang termasuk ke dalam virus ND genotipe VII dan VND yang bersirkulasi di Indonesia terbagi menjadi dua klaster besar.Diharapkan adanya penelitian lanjutan untuk mengetahui patogenisitas keempat VND isolat lapang dan uji tantang silang untuk formulasi bibit vaksin isolat lapang.
UCAPAN TERIMA KASIH
Ucapan terima kasih yang sebesar-besarnya kepada Indonesian Biodeversity Research Center (IBRC) Bali, atas bantuannya dalam penyediaan bahan dan data penelitian. Demikian pula, kepada teman-teman sejawat yang membantu penelitian ini, kami menyampaikan apresiasi.
DAFTAR PUSTAKA
Absalón AE., Mariano-Matías A, Vásquez-
Márquez A, Morales-Garzón A, Cortés-Espinosa DV, Ortega-García R dan Lucio-Decanini E. 2012. Complete genome sequence of a velogenic Newcastle disease virus isolated in Mexico. Virus Genes .45(2): 304-310.
ADHPI. 2011. Seminar ND Genotip 7B.
http://www.adhpi.org/seminar/seminar-nd-genotip-7b/.
Adi AAAM dan Astawa NM. 2014. Avian Paramyxovirus 1 : Biologi dan Polimorfisme Genetik. Swasta Nulus. hal 70.
Alexander DJ. 2000. Newcastle disease and other avian paramyxoviruses.Rev. sci. tech. Off. int. Epiz. 19 (2): 443-462.
Alexander DJ. 1987. Taxonomy and
Nomenclature of Avian Paramyxoviruses. Avian Patholog.16: 547-552.
Botus D. 2009. Predicting antigenic sites of hemagglutinin-neuraminidase glycoprotein – Newcastle disease virus.Romanian
Biotechnological Letters. 14(4):4589-4596.
Couceiro ESS, Couceiro JNSS dan Cabral MS.
1995. Hamegglutinating and Fusogenic Activities of the Newcastle Disease Virus : Studies on Receptor Binding Specifity and pH-induced Conformational Changes. Mern Inst Oswaldo Cruz.90(4):515-520.
Crennell S, Takimoto T, Portner A, dan Taylor G.
2000. Crystal structure of the multifunctional paramyxovirus hemagglutinin-
neuraminidas.Nature Structural & Molecular Biology.7: 1068 – 1074.
Dortmans JCFM. 2011.Virulence Determinants of Newcastle Disease Virus. Dissertation. Universitas Utrecht, Nederlands.
Dortmans JCFM, Koch G, Rottier PJM, Peeters BPH. 2011. Virulence of Newcastle disease virus: what is known so far?.Veterinary Research.42: 122.
Gong YY dan ZhiZhong CUI. 2011. Epitope variation in the Newcastle disease virus HN gene under antibody immune selective
pressure in cell culture.Sci China Life Sci. 54(5): 474–479.
Hughes AL. 2009. Relaxation of Purifying Selection on Live Attenuated VaccineStrains of the Family Paxamyxoviridae. J Vaccine, 27(11): 1685–1690.
Iorio RM, Syddall RJ, Sheehan JP, Bratt MA, Glickman RL, dan Riel AM. 1991. Neutralization Map of the HemagglutininNeuraminidase Glycoprotein of Newcastle Disease Virus: Domains Recognized by Monoclonal Antibodies That Prevent Receptor Recognition. Journal of Virology. 65(9): 4999-5006.
Iorio RM dan Bratt MA. 1984. Neutralization of Newcastle Disease Virus by Monoclonal Antibodies to the HemagglutininNeuraminidase Glycoprotein: Requirement for Antibodies to Four Sites for Complete Neutralization. Journal of Virology. 51(2):445-451.
Ke GM, Chuang KP, Chang CD, Lin MY, Liu HJ, 2010.Analysis of sequence and
haemagglutinin activity of the HN glycoprotein of Newcastle disease virus.Avian Pathology. 39(3): 235-244.
Kencana GAY, Kardena IM, dan Mahardika IGN. 2012. Peneguhan diagnosis penyakit Newcastle disease lapang pada ayam buras di Bali menggunakan teknik RT-PCR Jurnal Kedokteran Hewan. 6(1).
Khadzhiev G. 1987. Typing of Newcastle virus isolate by mean death time of chickens in the cloacal test.Vet Med NaukiI. 24(5):16-26.
Kusmaedi. 2001. Teknik uji hemaglutination inhibition Untuk mengukur tingkat kekebalan Terhadap newcastle disease dan egg drop Syndrome’76. Temu Teknis Fungsional Non Peneliti, Balai Penelitian Veteriner.
Mahon PJ, Mirza AM, Musich TA, dan Iorio RM. 2008. Engineered Intermonomeric Disulfide Bonds in the Globular Domain of Newcastle Disease Virus Hemagglutinin-Neuraminidase Protein: Implications for the Mechanism of Fusion Promotion.Journal of Virology. 83(21):10386–10396.
McGinnes LW dan Morrison TG. 2006. Inhibition of Receptor Binding Stabilizes Newcastle Disease Virus HN and F Protein-Containing Complexes. Journal of Virology, 80(6):
2894–2903.
Miller GL dan Stanley WM. 1944.Quantitative Aspects of the Red Blood Cell Agglutination Test for Influenza Virus. J Exp Med. 79(2): 185–195.
Oberdorfer AR, Werner O, Veits J, Mebatsion T, dan Mettenleiter TC. 2003. Contribution of the length of the HN protein and the sequence of the F protein cleavage site to Newcastle disease virus
pathogenicity.Journal of General Virology. 84: 3121–3129.
OIE. 2004. Newcastle Disease. Manual of diagnostic tests and vaccines for terrestrial animals, 2:1–15.
Panda A. 2003. "Role of Hemagglutinin
Neuraminidase Protein in Newcastle Disease Virus Pathogenesis” (dissertation).
University of Maryland.
Panda A, Elankumaran S, Krishnamurthy S, Huang Z, dan Samal SK. 2004. Loss of N-Linked Glycosylation from the
Hemagglutinin-Neuraminidase Protein Alters Virulenceof Newcastle Disease
Virus.Journal of Virology, 78 (10): 4965– 4975.
Perozo F, Marcano R, dan Afonsoc CL. 2012. Biological and Phylogenetic Characterization of a Genotype VII Newcastle Disease Virus from Venezuela: Efficacy of Field
Vaccination. Journal of Clinical
Microbiology.p:1204–1208.
Ponnusamy P, Kirubaharan JJ, dan Chandramohan A. 2009.Sequence Analysis of HN and F Genes of a less Virulent Newcastle Disease Virus Isolated from Unvaccinated Village Chicken. International Journal of Poultry Science. 8 (10): 985-994.
Saepulloh M dan Darminto. 2005. Kaman newcastle disease pada itik dan upaya Pengendaliannya WARTAZOA,Vol. 15 No . 2.
28
Discussion and feedback