KEMAMPUAN TANAH HUTAN MANGROVE SEBAGAI SUMBER ENZIM DALAM HIDROLISIS ENZIMATIK SUBSTRAT SEKAM PADI
on
ISSN 1907-9850
KEMAMPUAN TANAH HUTAN MANGROVE SEBAGAI SUMBER ENZIM DALAM HIDROLISIS ENZIMATIK SUBSTRAT SEKAM PADI
Ni Luh Md. Widayantini, I Nengah Wirajana*, dan Putu Suarya
Jurusan Kimia FMIPA Universitas Udayana, Bukit Jimbara *email : nwirajana@gmail.com
ABSTRAK
Aktivitas selulase tanah hutan mangrove dari Pantai Suwung Kauh, Bali telah dilaporkan pada penelitian sebelumnya. Telah dilakukan penelitian mengenai hidrolisis enzimatik sekam padi dengan menggunakan tanah hutan mangrove ini sebagai sumber enzim. Penelitian ini bertujuan untuk mengetahui kemampuan tanah hutan mangrove sebagai sumber enzim dalam menghidrolisis substrat sekam padi dengan dan tanpa delignifikasi yang diinkubasi dengan variasi waktu inkubasi tertentu. Substrat sekam delignifikasi dan tanpa delignifikasi masing-masing dicampur dengan tanah hutan mangrove lalu diinkubasi pada pH 7 dan suhu 29oC dengan variasi waktu inkubasi 0, 1, 2, 3, dan 4 minggu. Kadar gula pereduksi dari hasil inkubasi diukur secara spektrofotometri menggunakan metode Nelson-Somogyi. Hasil penelitian menunjukkan bahwa tanah hutan mangrove memiliki kemampuan dalam menghidrolisis sekam padi dengan delignifikasi, namun tidak mampu menghidrolisis sekam padi tanpa delignifikasi. Gula pereduksi tertinggi sebesar 0,829 mg/100mL dihasilkan dari hidrolisis substrat sekam delignifikasi pada minggu ke-1. Hasil ini mengindikasikan bahwa tanah hutan mangrove sebagai sumber selulase mempunyai kemampuan dalam menghidrolisis substrat sekam padi dengan delignifikasi pada kondisi pH 7 dan suhu 29oC dengan waktu inkubasi selama 1 minggu.
Kata kunci: sekam padi, delignifikasi, hidrolisis enzimatik, tanah hutan mangrove, gula pereduksi
ABSTRACT
Cellulase activity of mangrove forest soil from Suwung Kauh Beach in Bali has been reported in previous study. An enzymatic hydrolysis of rice husk with this mangrove forest soil as a source of enzymes was conducted. The aim of this study was to determine the ability of this mangrove forest soil as a source enzymes in hydrolyzing rice husk with and without delignification incubated in varied durations. The rice husk with and without delignification were mixed with mangrove forest soil and incubated at pH 7 and 29oC with a various incubation times of 0, 1, 2 , 3, and 4 weeks. Reducing sugar content of the results of incubation was measured by spectrophotometry using the Nelson-Somogyi method. The results of this study showed that the mangrove forest soil can hydrolyse the delignification rice husk, but could not degrade the rice husk without delignification. The highest reducing sugar content of 0,892 mg/100mL was resulted from hydrolysis of the delignification rice husk during one week. This result indicated that the mangrove forest soil as a source of cellulase had an ability in hydrolyzing the delignificated rice husk in pH 7 and 29oC incubated in one week.
Keywords: rice husk, delignification, enzymatic hydrolysis, mangrove forest soil, reducing suga
PENDAHULUAN
Kebutuhan manusia tehadap bahan bakar minyak (BBM) terus meningkat, sedangkan cadangan energi minyak bumi setiap harinya
semakin berkurang. Oleh karena itu, perlu dilakukan pengembangan sumber energi alternatif yang dapat diperbaharui dan murah untuk mengurangi ketergantungan terhadap BBM. Salah satu alternatif yang dapat menunjang kebutuhan
akan energi adalah dengan mengonversi biomassa menjadi bioetanol.
Produksi bioetanol melibatkan dua tahapan utama, yaitu hidrolisis dan fermentasi. Tahapan terpenting produksi bioetanol dari biomassa adalah tahapan hidrolisis untuk mendapatkan gula sederhana. Hasil fermentasi akan dipengaruhi oleh gula sederhana hasil hidrolisis karena gula sederhana itu yang nantinya akan difermentasi menjadi etanol (Anindyawati, 2009; Samsuri, et al., 2007).
Padi merupakan tanaman pangan yang keberadaannya melimpah di Indonesia dan ketersediaan sekam padi sebagai salah satu limbah penggilingan padi rata-rata sekitar 10 juta ton/tahun (Agung, et al., 2010). Pemanfaatan sekam padi umumnya hanya terbatas untuk campuran pupuk organik, media tanam holtikultura, peternakan ayam boiler, dan bahan bakar batu bata (Kurniawan dan Marsono, 2008). Sekam padi sebagai biomassa sebagian besar tersusun dari 31,12% selulosa; 22,48% hemiselulosa; dan 22,34% lignin (Kumar, et al., 2010). Bahan selulosa dan hemiselulosa dapat dimanfaatkan sebagai sumber karbon untuk produksi etanol dengan melakukan proses hidrolisis terlebih dahulu (Shofiyanto, 2008; Samsuri, et al., 2007).
Hidrolisis dapat dilakukan dengan metode asam dan enzimatik (Wyman, 1994). Hidrolisis dengan menggunakan asam tidak ramah lingkungan karena asam bersifat korosif, beracun, dan relatif mahal (Holm and Lassi, 2011; Wyman, 1994). Pengembangan teknologi bioproses dengan menggunakan enzim pada proses hidrolisisnya diyakini sebagai suatu proses yang lebih ramah lingkungan (Samsuri, et al., 2007). Beberapa penelitian melaporkan bahwa sekam padi dapat dimanfaatkan sebagai media pertumbuhan jamur penghasil enzim seperti Zymomonas mobilis, Lentinus polychous, dan Aspergillus niger (Rabah et al., 2011; Moonjai et al.,2007; Gunam, et al., 2010). Dengan kata lain, sekam padi dapat digunakan sebagai substrat untuk menghasilkan enzim. Hidrolisis biomassa secara enzimatik yang banyak dilakukan hanya menggunakan satu jenis enzim saja yang diisolasi dari mikroorganisme tertentu maupun enzim komersial yang harganya relatif mahal. Namun demikian, Widjaja dan Gunawan (2012) telah menggunakan campuran
enzim untuk menghidrolisis substrat jerami padi dan menunjukkan bahwa sistem campuran enzim dapat saling bekerja sama dalam degradasi komponen biomassa.
Tanah hutan mangrove merupakan ekosistem yang memiliki keanekaragaman mikroorganisme penghasil berbagai jenis enzim, salah satunya adalah enzim-enzim pendegradasi lignoselulosa. Tanah hutan mangrove di Bhitarkanika, Odisha, India dilaporkan memiliki aktivitas selulase. Sebanyak tujuh macam bakteri selulolitik telah ditemukan di sana, yaitu: 4 jenis Pseudomonas, dan masing-masing 1 jenis Bacillus polymyxa, B. mycoides, serta B. Brevis (Thatoi et al., 2012). Selain itu, Wirajana, et al. (2012) melaporkan adanya aktivitas selulase pada tanah hutan mangrove pantai Suwung Kauh, Bali dengan metode Filter Paper Assay dan Carboxymethyl Cellulose Assay. Pemanfaatan tanah secara langsung sebagai sumber enzim sangat menarik untuk dikaji di era green chemistry. Walaupun selama ini penggunaan mikroorganisme sebagai sumber enzim dalam industri dapat menekan biaya, namun penggunaan tanah sebagai sumber enzim yang lebih bervariatif dan tentunya jauh lebih murah akan menjadi tantangan dunia ilmu pengetahuan dan teknologi untuk memanfaatkannya.
Kandungan lignin yang berikatan kuat dengan selulosa dan hemiselulosa harus dihilangkan terlebih dahulu agar diperoleh selulosa dan hemiselulosa yang bebas dari lignin sehingga dapat dimanfaatkan lebih lanjut (Perez, et al., 2002). Penelitian Gunam, et al. (2010) melaporkan bahwa konsentrasi substrat jerami padi 2% dengan perlakuan delignifikasi menggunakan NaOH 6% menghasilkan aktivitas selulase yang optimal dari kapang Aspergillus niger. Penggunaan tanah hutan mangrove sebagai sumber berbagai enzim diduga mampu mendegradasi komponen lignoselulosa sekam padi yang kompleks. Maka dari itu, pada penelitian ini dilakukan perbedaan perlakuan terhadap substrat sekam padi yaitu dengan dan tanpa delignifikasi agar dapat digunakan sebagai pilihan dalam efisiensi waktu dan biaya jika menghasilkan kadar gula yang tidak berbeda jauh.
Kurniawati (2012) menggunakan tanah hutan mangrove Pantai Suwung Kauh, Bali sebagai sumber selulase yang aktivitasnya dapat ditingkatkan dengan substrat selulosa dari janur
kelapa. Hasil penelitian tersebut menyatakan bahwa janur kelapa diduga berperan sebagai penginduksi gen selulase dan aktivitas selulase dari tanah tersebut terus meningkat seiring dengan lamanya waktu inkubasi hingga 4 minggu. Aktivitas selulase yang meningkat mengindikasikan bahwa kemampuan mikroorganisme dalam menghasilkan enzim dan mengurai substrat juga meningkat. Berdasarkan penelitian tersebut, maka dilakukan hidrolisis enzimatik sekam padi dengan dan tanpa delignifikasi menggunakan tanah hutan mangrove sebagai sumber enzim dengan variasi waktu inkubasi 0, 1, 2, 3, dan 4 minggu.
MATERI DAN METODE
Bahan
Bahan yang digunakan dalam penelitian ini adalah sampel tanah dari konservasi hutan mangrove pantai Suwung Kauh-Bali diambil dari lokasi garis lintang 8o43’37,38”S dan garis bujur 115o11’42,10”T. Kondisi tanah menunjukkan suhu 29oC dan pH 7. Sekam padi diperoleh dari salah satu tempat penggilingan padi di Desa Nyitdah, Kabupaten Tabanan, Bali. Bahan-bahan kimia yang digunakan adalah bahan dengan kualitas proanalisis, antara lain: di-natrium hidrogenfosfat (Na2HPO4.2H2O), natrium di-hidrogenfosfat (NaH2PO4.H2O), glukosa anhidrat (C6H12O6), aquades (H2O), natrium hidroksida (NaOH), natrium karbonat anhidrat (Na2CO3), garam Rochelle (C4H4KNaO6.4H2O), natrium bikarbonat (NaHCO3), natrium sulfat anhidrat (Na2SO4), tembaga (II) sulfat pentahidrat (CuSO4.5H2O), asam sulfat pekat (H2SO4), ammonium molibdat tetrahidrat ((NH4)6Mo7O24.4H2O), di-natrium arsenat heptahidrat (Na2HAsO4.7H2O), aluminium foil, alkohol 70%, dan indikator universal.
Peralatan
Alat-alat yang digunakan di dalam penelitian ini adalah: ayakan, kaca arloji, pipet tetes, gelas piala (Beaker glass), Erlenmeyer, labu ukur, tabung reaksi, kuvet, batang pengaduk, botol semprot, spatula, pipet volume,TenSette pipette, tip, termometer, neraca analitik, inkubator, autoclave, hot plate, alat sentrifugasi, dan
Spektrofotometer UV-Vis 1800 Shimadzu Double Beam.
Cara Kerja
Pengambilan sampel tanah hutan mangrove
Sampel tanah basah diambil dari lokasi garis lintang 8o43’37,38”S dan garis bujur 115o11’42,10”T di kawasan hutan mangrove Pantai Suwung Kauh, Bali yang banyak terdapat daun-daun (limbah organik) yang telah membusuk. Hal ini ditujukan untuk meyakinkan adanya kemungkinan aktivitas mikroba pendegradasi biomassa. Pengambilan tanah dilakukan pada area sekitar 5 m2 yang diambil di lima titik secara acak. Tanah hutan mangrove diukur pH dan suhunya di kedalaman 0-10 cm dari permukaan tanah. Setelah pH dan suhunya dicatat, tanah basah dari hutan mangrove diambil di kedalaman 0 – 10 cm lalu tanah di lima titik tersebut dicampur menjadi satu dalam wadah polietilen steril.Tanah dapat disimpan dalam kondisi dingin (kulkas) sebelum digunakan untuk langkah berikutnya.
Persiapan substrat sekam padi Perlakuan fisik
Sekam padi dicuci terlebih dahulu lalu dikeringkan. Sekam padi yang telah kering selanjutnya diblender kemudian diayak dengan ayakan 150 µm dan 75 µm sehingga diperoleh serbuk sekam padi ukuran 75-150 µm.
Delignifikasi sekam padi
Sebanyak 10 gram serbuk sekam padi direndam dengan NaOH 6% (b/v) dalam
Erlenmeyer 250 mL dengan perbandingan 1:5 (serbuk sekam padi:larutan NaOH) selanjutnya dipanaskan dengan autoclave pada suhu 121oC selama 15 menit lalu disaring dengan kain saring. Serbuk yang telah disaring selanjutnya dicuci dengan aquades hingga netral (pH = 7) lalu
dikeringkan dengan oven pada suhu 105oC selama 10 jam (Gunam dkk., 2010). Sekam padi yang mengalami delignifikasi selanjutnya diberi nama substrat sekam D.
Sekam padi tanpa delignifikasi
Sekam padi yang telah mengalami perlakuan fisik selanjutnya siap digunakan tanpa diproses dengan delignifikasi. Sekam padi ini selanjutnya diberi nama substrat sekam TD.
Inkubasi substrat sekam padi dengan tanah hutan mangrove
Inkubasi sekam padi dengan tanah hutan mangrove dilakukan berdasarkan adopsi dan modifikasi metode yang dilakukan Kurniawati (2012) dan Wirajana, et al. (2012). Inkubasi dilakukan dalam Erlenmeyer 100 mL. Substrat sekam D dan TD masing-masing dimasukkan ke dalam Erlenmeyer sebanyak 0,10 gram lalu disterilisasi pada suhu 121oC selama 15 menit dengan autoclave. Selanjutnya tanah hutan mangrove ditambahkan sebanyak 30,00 g, lalu ditambahi 3,0 mL bufer fosfat kemudian diaduk agar merata. Sampel uji diinkubasi dengan variasi waktu 0, 1, 2, 3, dan 4 minggu pada suhu 29oC di dalam inkubator.
Analisis Gula Pereduksi Hasil Inkubasi
Hasil inkubasi antara substrat sekam dengan tanah hutan mangrove diambil sebanyak 10 gram dan disentrifugasi pada kecepatan 3000 rpm (800 xg) selama 15 menit untuk mendapatkan filtrat yang jernih. Filtrat lalu diukur kadar gula pereduksinya dengan metode Nelson-Somogyi. (Sudarmadji, et al., 1997).
HASIL DAN PEMBAHASAN
Perlakuan Awal terhadap Sekam Padi
Perlakuan fisik terhadap sekam padi dilakukan dengan memperkecil ukuran sekam padi menggunakan blender kemudian diayak dengan ayakan 75 µm dan 150 µm sehingga diperoleh sekam padi dengan ukuran partikel 75-150 µm. Semakin kecil ukuran partikel substrat, maka luas permukaan kontak antara substrat dan enzim akan semakin besar atau mudah (Mosier, et al., 2005). Sekam padi yang telah diayak selanjutnya disterilisasi dengan autoclave pada suhu 120oC selama 15 menit. Sterilisasi ini dimaksudkan untuk mematikan mikroorganisme yang ada pada sekam padi agar meyakinkan pada proses inkubasi dengan tanah hutan mangrove, mikroorganisme penghasil enzim untuk mendegradasi poliskarida sekam padi hanya berasal dari tanah hutan mangrove saja.
Delignifikasi dilakukan untuk menghilangkan kandungan lignin dalam sekam padi. Hal tersebut dilakukan karena polisakarida (selulosa dan hemiselulosa) yang terdapat pada
sekam padi tidak berada dalam keadaan bebas melainkan selalu terikat sangat kuat dengan lignin. Keadaan yang demikian dapat mengakibatkan enzim akan sulit mengakses selulosa dan hemiselulosa.
Menurut Safaria, et al. (2013), ion OH-dari NaOH akan memutuskan ikatan-ikatan dari struktur dasar lignin sedangkan ion Na+ akan berikatan dengan lignin membentuk natrium fenolat. Garam fenolat ini bersifat mudah larut dalam air karena sifat kepolarannya. Lignin yang terlarut ditandai dengan warna hitam pada larutan yang disebut lindi hitam (black liquor).
Hasil Analisis Gula Pereduksi
Substrat sekam D dan substrat sekam TD masing-masing diinkubasi bersama dengan tanah hutan mangrove sebagai sumber enzim. Pada prinsipnya, dengan kehadiran substrat, mikroorganisme yang terdapat di tanah hutan mangrove akan memanfaatkan substrat tersebut sebagai makanannya dan menghasilkan enzim untuk mengurai substrat tersebut dalam proses metabolismenya. Produk yang diharapkan dari degradasi sekam padi adalah berupa gula sederhana penyusun sekam padi, yaitu glukosa sebagai monomer selulosa serta xilosa, arabinosa, dan manosa sebagai monomer utama penyusun hemiselulosa (Howard et al., 2003) yang terukur sebagai gula pereduksi menggunakan metode Nelson-Somogyi. Absorbansi larutan standar dan larutan sampel diukur dengan spektrofotometer UV-Vis pada panjang gelombang 540 nm dengan menggunakan metode standar tunggal.
Absorbansi yang diperoleh dikonversi ke kadar gula pereduksi mg/100mL sesuai dengan rumus berikut (Skoog, 1985) :
C
sampel
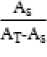
x C
standar
Keterangan :
Csampel : konsentrasi sampel (mg/100mL)
Cstandar : konsentrasi standar (mg/100mL)
As : absorbansi sampel
AT : absorbansi total (larutan standar dan
sampel)
Pada tanah hutan mangrove kemungkinan terdapat gula sederhana yang tidak berkaitan dengan hidrolisis enzimatik substrat oleh tanah
hutan mangrove sebagai sumber enzim. Hal ini disebabkan oleh nutrien gula yang memang sudah ada di dalam sampel tanah saat pengambilan dan dapat terukur pada saat analisis sebagai gula pereduksi. Hal ini dapat membiaskan jumlah gula pereduksi yang sebenarnya dihasilkan dalam hidrolisis sekam oleh enzim yang ada di tanah Oleh karena itu, maka dibuatlah kontrol ditentukan langsung kadar gula pereduksi pada saat t=0.
Profil hubungan antara lamanya waktu inkubasi dengan kadar gula pereduksi disajikan secara deskriptif oleh grafik pada Gambar 1.
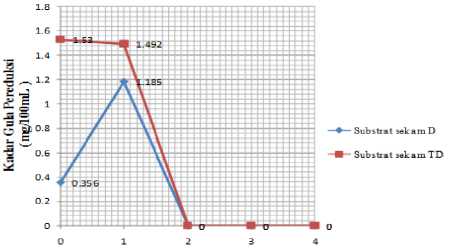
Wakni Inkubasi ( minggu )
Gambar 1. Pengaruh waktu inkubasi terhadap kadar gula pereduksi
Inkubasi substrat sekam D dengan tanah hutan mangrove sebagai sumber enzim selama 1 minggu menunjukkan kadar gula pereduksi yang terukur sebesar 1,185 mg/100mL, dimana pada t=0 terukur pula gula pereduksi sebesar 0,356 mg/100mL. Dengan demikian, selama selang waktu 1 minggu telah terbentuk gula pereduksi sebanyak 0,829 mg/100mL. Akan tetapi, setelah selang waktu 2 minggu, 3 minggu, hingga 4 minggu pengukuran terhadap gula pereduksi hasil inkubasi tidak terdeteksi.
Terbentukya gula pereduksi dari hasil inkubasi substrat sekam D dengan tanah hutan mangrove mengindikasikan bahwa telah terjadi penguraian substrat oleh mikroorganisme penghasil enzim. Kadar gula yang tidak terdeteksi diduga disebabkan oleh adanya aktivitas mikroorganisme dalam sampel tanah yang semakin tinggi dalam memanfaatkan gula sebagai sumber karbon untuk metabolismenya. Tingkat konsumsi gula oleh mikroorganisme yang ada dalam sampel tanah lebih tinggi dibandingkan dengan kecepatan hidrolisis yang menyebabkan terjadinya penurunan kadar gula pereduksi.
Interaksi antara enzim dan substrat yang semakin lama menyebabkan reaksi berjalan lebih maksimal sehingga konsentrasi gula yang dihasilkan menjadi lebih tinggi. Akan tetapi pada waktu hidrolisis tertentu konsentrasi gula akan mengalami penurunan (Safaria, et al., 2013). Setelah sumber karbon dalam media tumbuhnya hampir habis, maka mikroorganisme akan memanfaatkan kembali gula sederhana yang telah diuraikan sebagai sumber energi dalam selnya, yang ditandai dengan menurunnya kadar gula pereduksi yang dihasilkan. Hal tersebut didukung oleh penelitian Safaria, et al. (2013) yang menghidrolisis substrat sabut kelapa menggunakan selulase dari Aspergillus niger dan Trichoderma reesei dimana kadar gula pereduksi maksimal dicapai pada waktu interaksi enzim dengan substrat 6 jam dan akhirnya terus-menerus menurun pada jam ke-7 dan ke-8. Begitu pula dengan penelitian Dewi, e al. (2005) mengenai produksi gula pereduksi oleh Rhizopus oryzae dari substart bekatul memperoleh kadar gula pereduksi maksimal pada waktu interaksi enzim dengan substrat 1 hari dan terus-menerus mengalami penurunan pada hari ke-2 dan ke-3 akibat konsumsi gula pereduksi semakin meningkat oleh R. oryzae yang mulai bersporulasi sehingga gula pereduksi dikonsumsi untuk proses sporulasi.
Pemanfaatan nutrien gula sederhana oleh mikroorganisme yang tersedia di media inkubasi dikuatkan pula oleh hasil inkubasi substrat sekam TD. Hasil pengukuran terhadap gula pereduksi yang terus menerus mengalami penurunan mengindikasikan bahwa tidak terjadinya proses hidrolisis substrat sekam TD. Terukurnya gula pereduksi pada t=0 menandakan bahwa kandungan gula yang memang sudah ada di tanah pada saat pengambilan sampel. Kadar gula pereduksi yang terukur tiap minggu juga berasal dari sampel tanah yang terbentuk di lingkungan tanah tersebut diambil dan terus berkurang karena dimanfaatkan oleh mikroorganisme yang ada pada sampel tanah.
Hasil tersebut secara deskriptif membuktikan bahwa adanya pengaruh perlakuan awal delignifikasi terhadap substrat sekam padi. Selulosa maupun hemiselulosa pada substrat sekam TD masih terikat kuat oleh lignin yang menghambat akses enzim untuk menerobos perisai lignin sehingga penguraian selulosa dan hemiselulosa terhambat dan tidak ada gula
sederhana yang dihasilkan. Fenomena tersebut juga dapat mengindikasikan bahwa di dalam sampel tanah yang diambil hampir tidak ada mikroorganisme yang mampu mendegradasi lignin. Akan tetapi, untuk memastikan hal itu perlu dilakuan penelitian lebih lanjut untuk mengidentifikasi mikroorganisme yang ada di ekosistem tanah hutan mangrove pantai Suwung Kauh-Bali, mengingat masih belum ada peneliti yang melakukannya. Menurut Perez et al. (2002) terdapat kapang pelapuk putih jenis P. chrysosporium merupakan salah satu kapang yang efektif dan sering dijadikan model dalam pengujian degradasi komponen lignoselulosa terutama lignin. Kurniawati (2012) melaporkan bahwa aktivitas selulase tanah hutan mangrove dengan pengayaan selulosa janur kelapa mengalami peningkatan terus menerus dengan lama pengayaan selama 4 minggu. Namun demikian, dari hasil penelitian ini ternyata aktivitas enzim yang meningkat tidak menjamin produk hasil degradasi substrat oleh enzim juga akan meningkat. Hal ini disebabkan oleh
beranekaragamnya komunitas mikroorganisme
yang hidup di tanah hutan mangrove sehingga produk hasil degradasi substrat dimanfaatkan pula oleh mikroorganisme lain dalam sampel tanah maupun mikroorganisme pendegradasi selulosa dan hemiselulosa itu sendiri saat sumber karbon dalam media inkubasi mulai habis.
SIMPULAN DAN SARAN
Simpulan
Tanah hutan mangrove pantai Suwung Kauh, Bali memiliki kemampuan sebagai sumber enzim dalam mendegradasi substrat sekam padi delignifikasi. Inkubasi substrat sekam tanpa delignifikasi tidak dapat menghasilkan gula pereduksi. Waktu inkubasi antara substrat sekam padi dengan tanah hutan mangrove sebagai sumber enzim mempengaruhi kadar gula pereduksi yang dihasilkan. Gula pereduksi tertinggi sebesar 0,829 mg/100mL dihasilkan dari inkubasi substrat sekam delignifikasi selama 1 minggu. Waktu inkubasi yang lebih lama lagi, yakni sampai 4 minggu menyebabkan gula pereduksi berkurang terus.
Saran
Perlu dilakukan pengembangan penelitian mengenai metode yang dapat digunakan secara tepat dalam mengekstraksi dan analisis gula pereduksi yang diperoleh dari suspensi tanah.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Kepala Laboratorium Sumber Daya Genetika dan Biologi Molekuler Gedung Pascasarjana Universitas Udayana dan Kepala Laboratorium Bersama FMIPA Universitas Udayana yang membantu dalam pelaksanaan penelitian serta Ir. Wahyu Dwijani, M.Kes. dan Drs. I Wayan Suarsa, M.Si. yang memberikan kritik dan saran dalam penulisan hasil penelitian ini.
DAFTAR PUSTAKA
Agung, W.W., Pranolo, S.H., Noorochadi, G., dan Ratna M., L., 2010, Perancangan dan Uji-Kinerja Reaktor Gasifikasi Sekam Padi Skala Kecil, Ekuilibrium, 9 (1) : 29-33
Anindyawati, T., 2009, Prospek Enzim dan
Limbah Lignoselulosa untuk Produksi Bioetanol, BS., 44 (1) : 49-56
Dewi, C., Purwoko, T., dan Pangastuti, A., 2005, Produksi Gula Reduksi oleh Rhizopus oryzae dari Substrat Bekatul, Bioteknologi, 2 (1) : 21-26
Gunam, I.B.W., Buda, K., dan Guna, I.M.Y.S., 2010, Pengaruh Perlakuan Delignifikasi dengan Larutan NaOH dan Konsentrasi Substrat Jerami Padi terhadap Produksi Enzim Selulase dari Aspergillus niger NRRL A-II, 264, Jurnal Biologi, XIV (1) : 55-61
Holm, J. and Lassi, U., 2011, Ionic Liquids in The Pretreatment of Lignocellulosic Biomass, Ionic Liquids: Applications and
Perspectives, Prof. Alexander Kokorin (Ed.), ISBN: 978-953-307-248-7, InTech, 545-560
Howard, R.L., E., Abotsi, Jansen van Rensburg E.L., and Howard S., 2003,
Lignocellulose Biotechnology: Issues of Bioconversion and Enzyme
Production:Review, African Journal of Biotechnology, 2 (12) : 602-619
Kumar, P.S., Ramakrishnan, K., Kirupha, S.D., and Sivanesan, S., 2010, Thermodynamic and Kinetic Studies of Cadmium Adsorption from Aqueous Solution onto Rice Husk, Brazilian Journal of Chemical Engineering, 27 (02) : 347 – 355
Kurniawan, O. dan Marsono, 2008, Superkarbon Bahan Bakar Alternatif, Niaga Swadaya, Jakarta
Kurniawati, I., 2012, Aktivitas Enzim Selulase dari Tanah Hutan Mangrove Pantai Suwung Bali dengan Pengayaan Substrat Janur Kelapa (Cocus nucifera), Tesis,
Universitas Udayana, Denpasar
Moonjai, N., Pukahuta, C., and Salubchua J., 2007, Simultaneous Saccharification and Fermentation of Fungal Biopretreated Rice Husk and Rice Polish to Ethanol, POSTER, 2-5
Mosier, N., Wyman, C., Dale, B., Elander, R., Lee, Y.Y., Holtzapple, M., and Ladisch, M., 2005, Features of Promising Technologies for Pretreatment of Lignocellulosic Biomass, Bioresource Technology, 96 :
673-686
Perez J., Munoz-Dorado, J., de la Rubia, T. and Martinez, J., 2002, Biodegradation and Biological Treatments of Cellulose, Hemicellulose, and Lignin: An Overview. Int. Microbiol, 5 : 53-63
Rabah, A.B., Oyeleke S. B., Manga S. B., and Hassan, L.G., 2011, Microbial
Pretreatment of Rice Husk and Groundnut Shell for Bioethanol Production, Int. Res. J. Microbiol, 2 (8) : 253-258
Safaria, S., Idiawati, N., dan Zaharah, T.A., 2013, Efektivitas Campuran Enzim Selulase dari
Aspergillus niger dan Trichoderma reesei dalam Menghidrolisis Substrat Sabut Kelapa, JKK, 2 (1) : 46-51
Samsuri, M., Gozan, M., Mardias, R., Baiquni, M., Hermansyah, H., Wijanarko, A., Prasetya, B., dan Nasikin, M., 2007, Pemanfaatan Selulosa Bagas untuk Produksi Ethanol Melalui Sakarifikasi dan Fermentasi Serentak dengan Enzim Xylanase, Makara Teknologi, 11 (1) : 17-24
Shofiyanto, M. E., 2008, Hidrolisis Tongkol
Jagung oleh Bakteri Selulolitik untuk Produksi Bioetanol dalam Kultur Campuran, Skripsi, Institut Pertanian Bogor, Bogor
Skoog, D.A., 1985, Principles of Instrumental
Analysis, 3rd edition, Saunders College
Publishing, Tokyo
Sudarmadji, S., Haryono, B., dan Suhardi, 1997, Prosedur Analisa untuk Makanan dan Pertanian, Edisi Keempat, Liberty, Yogyakarta
Thatoi, H., Behera, B.C., Dangar, T.K., and Mishra, R.R., 2012, Microbial Biodiversity in Mangrove Soils of Bhitarkanika, Odisha, India, International Journal of Environmental Biology, 2 (2) : 50-58
Widjaja, A. dan Gunawan S., 2012, Teknologi Produksi Bioetanol Generasi 2 Melalui Pemanfaatan Selulosa dan Hemiselulosa dalam Jerami Padi, Prosiding:InSINas, Surabaya, 1-6
Wirajana, I. N., Ratnayani, K., dan Yuliana, D. A., 2012, Skrining Selulase dan Isolasi DNA Metagenomik dari Tanah Hutan Mangrove Pantai Suwung Bali, Jurnal Kimia, 6 (2) : 191-195
Wyman, C. E., 1994, Ethanol from Lignocellulosic Biomass:Technology, Economics, and Opportunities, Bioresource Technology, 50 : 3-16
41
Discussion and feedback