PROFILE IMMUNOGLOBULIN M (IgM) BALI CATTLE IN NUSA PENIDA KLUNGKUNG DISTRICT BALI PROVINCE
on
Volume 11 No. 1: 85-93
Pebruari 2019
DOI: 10.24843/bulvet.2019.v11.i01.p14
Buletin Veteriner Udayana
pISSN: 2085-2495; eISSN: 2477-2712
Online pada: http://ojs.unud.ac.id/index.php/buletinvet
Terakreditasi Nasional Peringkat 3, DJPRP Kementerian Ristekdikti No. 21/E/KPT/2018, Tanggal 9 Juli 2018
Profil Imunoglobulin M Sapi Bali di Pulau Nusa Penida Klungkung Bali
(PROFILE IMMUNOGLOBULIN M (IgM) BALI CATTLE IN NUSA PENIDA ISLAND KLUNGKUNG DISTRICT BALI PROVINCE)
Luh Kadek Nanda Laksmi1*, I Nengah Kerta Besung2, I Nyoman Suartha3, Ni Ketut Suwiti4
-
1Balai Besar Veteriner Denpasar, Jl. Raya Sesetan No.266, Denpasar Selatan, Denpasar, Bali. 2Laboratorium Mikrobiologi Veteriner, Fakultas Kedokteran Hewan, Universitas Udayana.
-
3Laboratorium Ilmu Penyakit Dalam, Fakultas Kedokteran Hewan, Universitas Udayana. 4Departemen Histologi, Fakultas Kedokteran Hewan, Universitas Udayana.
*Email: laksmidvmnanda@gmail.com
ABSTRAK
Penelitian ini bertujuan untuk mengetahui profil imunoglobulin M (IgM) sapi bali di Nusa Penida. Sampel berupa serum dari 54 ekor sapi bali yang dipelihara di Nusa Penida berdasarkan kriteria geografis wilayah terdiri dari dataran tinggi (Desa Klumpu dan Batumadeg) dan dataran rendah (Desa Ped), umur meliputi sapi bali muda (12-18 bulan) dan dewasa (24 bulan ke atas), sementara jenis kelamin (jantan dan betina). Serum yang diperoleh diuji dengan menggunakan metode ELISA (Enzyme Linked Immunosorbent Assay). Hasil menunjukkan bahwa 54 sampel serum sapi bali di Nusa Penida terdeteksi adanya kadar imunoglobulin M (IgM) dengan nilai yang bervariasi berkisar dari 2.565 ng/mL – 8.834 ng/mL. Rerata kadar IgM serum sapi bali yang di pelihara di dataran rendah (4.837±1.385 ng/mL) secara deskriptif lebih tinggi daripada di dataran tinggi (4.761±1.353 ng/mL), sementara untuk sapi bali betina (5.018±1.370 ng/mL) lebih tinggi dibandingkan sapi bali jantan (4.477±1.290 ng/mL), sedangkan sapi bali dewasa (4.869±1.417ng/mL) lebih tinggi daripada sapi bali muda (4.707±1.305 ng/mL), akan tetapi secara statistik perbandingan dari semua kategori menujukkan tidak berbeda nyata (P>0.05). Rerata kadar IgM sapi bali yang dipelihara di Nusa Penida dapat di gunakan sebagai data base profil IgM sapi bali yang dipelihara di Nusa Penida.
Kata kunci: Imunoglobulin M; sapi bali; Nusa Penida; geografis; jenis kelamin; umur; ELISA.
ABSTRACT
This study aims to determine the profile of immunoglobulin M (IgM) of bali cattle in Nusa Penida. The serum samples of 54 bali cattle that were kept in Nusa Penida based on geographical criteria consisted of highlands (Klumpu and Batumadeg villages) and lowlands (Ped villages), age covering young bali cattle (12-18 months) and adults (24 months and over), while sex (male and female). The serum obtained was tested using the ELISA (Enzyme Linked Immunosorbent Assay) method. The results showed that 54 samples of serum of bali cattle in Nusa Penida were detected in the presence of levels of immunoglobulin M (IgM) with values ranging from 2.565 ng/mL – 8.834 ng/mL. The mean rate of serum IgM cattle raised in the lowlands (4,837 ± 1,385 ng / mL) was descriptively higher than in the highlands (4,761 ± 1,353 ng / mL), while for female bali cattle (5.018 ± 1.370 ng / mL) higher than male (4,477 ± 1.290 ng / mL), while adult bali cattle (4,869 ± 1.417 ng / mL) was higher than young bali cattle (4,707 ± 1.305 ng / mL), but statistically comparable from all categories showed no significantly (P> 0.05). The average rate of IgM of bali cattle maintained in Nusa Penida can be used as a data base of IgM profile of bali cattle that is maintained in Nusa Penida.
Keywords: Immunoglobulin M; bali cattle; Nusa Penida; geographic; gender; age; ELISA.
PENDAHULUAN
Sapi bali merupakan plasma nuftah asli Indonesia yang didomestikasi di Pulau Bali, Permentan No. 36 Tahun 2006 (Departemen Pertanian, 2006). Nusa Penida menjadi salah satu tempat pemurnian dan pembibitan sapi bali yang ditetapkan oleh Dirjen Peternakan dan Kesehatan Hewan pada tahun 2002. Sapi bali yang dipelihara di Nusa Penida telah diketahui secara subklinis bebas penyakit Jembrana dan SE (Septicemia Epizootica) (Suwiti et al., 2009).
Cekaman yang tinggi menyebabkan terjadinya stres. Kondisi ini dapat menginduksi perubahan fungsi imunitas selular dan humoral di dalam tubuh. Perubahan fungsi imunitas ini memberikan kerentanan terhadap suatu penyakit (Carrol dan Forsberg, 2007). Sapi bali rentan terhadap penyakit jembrana, ingusan (malignat catarrhal fever) dan bali ziekte (Damayanti, 2016; Indriawati et al., 2013). Sapi bali juga sering terinfestasi penyakit parasit (Antara et al., 2017; Indraswari et al., 2017). Adanya penyakit yang menyerang sapi bali akan berakibat menurunnya produktivitas, menambah biaya pengobatan, dan bahkan menimbulkan kematian.
Ketahanan tubuh sapi bali dapat diketahui melalui penilaian respon imun di dalam tubuh (Putro, 2004). Indikator ketahanan tubuh dapat dideteksi dari respon imun melalui pengukuran produksi imunoglobulin M (IgM). IgM merupakan kelas imunoglobulin yang awal dibentuk ketika terjadi rangsangan antigen. IgM ini berperan mencegah gerakan mikroorganisme patogen, memudahkan fagositosis, aglutinator poten protein, sebagai reseptor permukaan sel B, dan untuk perlekatan antigen. IgM memiliki kelebihan berupa efisiensi reaksi aglutinasi dan reaksi sitolitik sehingga mampu di produksi sangat cepat setelah infeksi dan tetap berada dalam darah (Abbas et al., 2007; Kresno, 2001).
Adanya IgM di dalam tubuh dipengaruhi oleh faktor umur dan jenis
kelamin (Surhayati dan Hartono, 2015; Telupere et al., 2014; Jazek et al., 2012; Iskandar, 2011; Rasyid et al., 2008). Faktor yang lain seperti letak geografis wilayah pemeliharaan di dataran tinggi dan rendah, ketiadaan sumber pakan dan cekaman suhu akan berpengaruh terhadap kesehatan sapi. Kondisi ini juga secara tidak langsung berdampak pada kadar IgM di dalam tubuh. Deteksi IgM di dalam tubuh penting dilakukan sebagai upaya untuk deteksi kejadian penyakit. Namun sampai saat ini belum ada studi tentang profil IgM pada sapi bali, sehingga data awal standar kadar IgM pada berbagai aspek seperti ketinggian tempat yang berbeda, tingkat umur, dan jenis kelamin belum diketahui secara pasti.
MATERI DAN METODE
Sampel Penelitian
Sampel yang digunakan dalam penelitian eksploratif eksperimental untuk menegetahui profil imunoglobulin M (IgM) adalah lima puluh empat (54) serum sapi bali yang dipelihara di Nusa Penida berdasarkan kriteria geografis wilayah terdiri dari dataran tinggi (desa Klumpu dan Batumadeg) dan dataran rendah (desa Ped), umur meliputi sapi bali muda (12-18 bulan) dan dewasa (24 bulan ke atas), sementara jenis kelamin (jantan dan betina).
Enzyme Linked Immunosorbent Assay
Uji ELISA yang dipergunakan dalam penelitian ini mengacu pada metode Jazek et al., (2012) dan Bayram et al., (2016) dengan beberapa modifikasi. Standard diencerkan dengan cara mencampurkan standard dengan standart dilution. Pertama-tama disiapkan 5 tabung efendorf, kemudian diambil 50 μl standart dilution untuk diisikan pada setiap tabung efendorf, selanjutnya diambil sebanyak 100 μl standart (360 ng/ml) dan dimasukkan pada tabung yang pertama. Sebanyak 100 μl larutan (campuran standart dilution dan standart) diambil dari tabung pertama dan dimasukkan ke dalam tabung ke dua. Sebanyak 50 μl larutan dari tabung yang kedua diambil dan dimasukkan ke dalam tabung yang ketiga. Pada tahap ketiga ini
diulang hingga tabung kelima sehingga konsentrasi standart senilai 20 ng/ml.
Pengujian pertama kali dengan disiapkan well untuk blank dan sampel, akan tetapi pada well blank tidak dilakukan penambahan sampel dan Horseradish Peroxidase (HRP) – Conjugate. Selanjutnya dimasukkan 40 μl sampel dilution ke masing-masing well dan ditambahkan 10 μl sampel yang di uji pada plate uji. Setelah itu di inkubasi dengan ditutup menggunakan adhesive strip selama 30 menit pada suhu 37oC. Pada saat pengenceran wash solution digunakan perbandingan 1: 20 dan wash solution diencerkan 30 kali lipat dengan menggukan destilled water. Hasil wash solution dipakai pada saat proses washing. Pada saat washing, strip adhesive dibuka, larutan yang ada di well di hilangkan, kemudian tambahkan washing buffer pada setiap well selama 30 detik, setelah itu dikeringkan. Proses washing dilakukan sebanyak 5 kali. Selanjutnya ditambahkan enzim HRP-Conjugate reagent 50 μl pada setiap well kecuali well blank. Kemudian diinkubasi menggunakan adhesive strip selama 30 menit pada suhu 37oC. Setelah 30 menit, dilakukan washing selama 30 detik sebanyak 5 kali pengulangan. Setelah proses washing dilanjutkan dengan pewarnaan dengan cara ditambahkan Chromogen Solution A dan Chromogen Solution B 50 μl pada setiap well selama 15 menit pada suhu 37oC. Setelah 15 menit dilakukan stop reaction dengan cara ditambahkan stop solution 50 μl pada setiap well, pada proses stop reaction akan terjadi perubahan warna dari warna biru menjadi warna kuning dan ditunggu selama 15 menit. Pada saat mendekati 24 menit ke 15 dilakukan pembacaan absorbance (daya serap) menggunakan ELISA reader 450 nm dengan blank well bernilai nol. Konsentrasi IgM pada sampel ditentukan dengan membandingkan nilai O.D dari sampel dengan kurva standar. Pada penilaian ini konsentrasi standart diibaratkan sebagai bidang horisontal, sedangkan nilai O.D
pada bidang vertikal. Gambar kurva standar pada kertas grafik. Konsentrasi yang bersesuaian dicari berdasarkan nilai dari O.D sampel dengan kurva sampel dan diperoleh dengan menggunakan persamaan X= aY*b-0.5, dengan X adalah konsentrasi IgM dalam serum sapi bali, Y adalah nilai O.D, adalah konstatnta dan b adalah koefisien. Perbedaan rerata kadar IgM serum sapi bali dianalisis dengan uji Independent T-test dengan menggunakan program software SPSS 17 (Sampurna dan Nindhia, 2008; Sampurna, 2012).
HASIL DAN PEMBAHASAN
Kadar imunoglobulin M (IgM) hasil uji terhadap serum sapi bali yang dipelihara di Nusa Penida Kabupaten Klungkung Provinsi Bali menggunakan metode Enzyme Linked Immunosorbent Assay (ELISA) dengan Bovine IgM ELISA kit (Glory Science Co., Ltd., Catalog: 14543) telah diperoleh nilai optical density (OD) berkisar dari 0,055-2,886. Hasil nilai optical density yang selanjutnya digunakan untuk menentukan kadar imunoglobulin M (IgM) pada serum sapi bali berdasarkan persamaan X= 53.747Y*1.579-0.5 dengan nilai (R2 = 0.975). Sebanyak 54 sampel serum sapi bali di Nusa Penida yang diuji telah diperoleh bahwa semua sampel serum sapi bali terdeteksi adanya kadar IgM yang bervariasi dengan kisaran 2.565 ng/mL – 8.834 ng/mL.
Kadar IgM serum sapi bali di dataran tinggi memiliki nilai bervariasi berkisar antara 2.565 ng/mL - 8.834 ng/mL, sedangkan kadar IgM serum sapi bali di dataran rendah memiliki nilai bervariasi berkisar antara 2.929 ng/mL - 8.217 ng/mL. Kadar IgM serum sapi bali betina memiliki nilai bervariasi berkisar antara 2.775 ng/mL - 8.834 ng/mL, sementara kadar IgM serum sapi bali jantan memiliki nilai bervariasi juga berkisar antara 2.565 ng/mL - 6.742 ng/mL. Kadar IgM serum sapi bali muda memiliki nilai bervariasi berkisar antara 2.775 ng/mL - 8.217 ng/mL, sedangkan kadar IgM serum sapi bali dewasa memiliki nilai bervariasi juga berkisar antara 2.565
ng/mL - 8.834 ng/mL. Rerarta kadar IgM serum sapi bali pada setiap kategori yaitu geografis wilayah, jenis kelamin, dan umur terlihat pada Grafik 1.
Terdeteksinya kadar IgM dalam serum pada penelitian ini menujukkan bahwa sapi
bali yang dipelihara di Nusa Penida telah menujukkan respon yang baik oleh innate IgM ataupun adaptive IgM terhadap antigen.
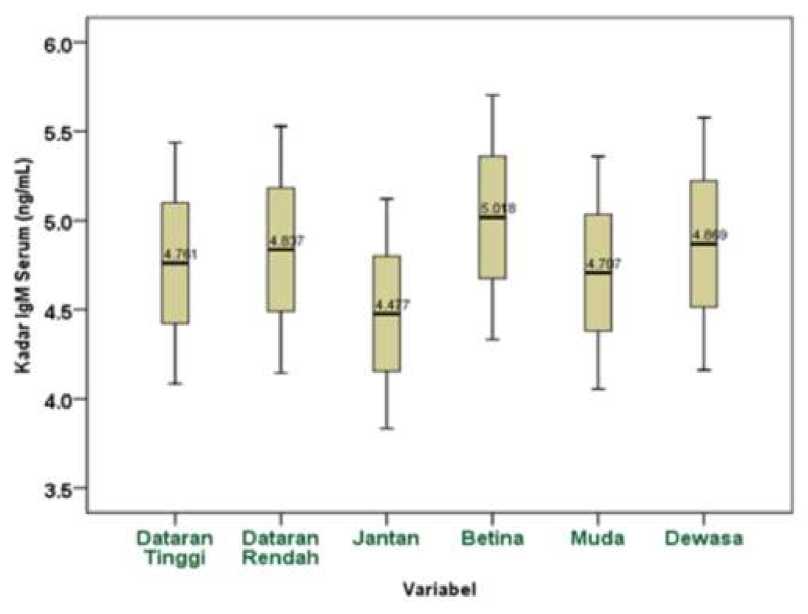
Grafik 1. Grafik Rerata Kadar Imunoglobulin M (IgM) Sapi Bali Berdasarkan Kriteria Geografis Wilayah (Dataran Tinggi dan Dataran Rendah), Jenis Kelamin (Jantan dan Betina), Umur (Muda dan Dewasa).
Pada studi sebelumnya telah diketahui kadar IgM pada susu sapi dengan kondisi sapi yang sehat yaitu 0,12 g/L (Korhonen et al., 1995) dan 1,23-1,65 g/L (Zagorska et al., 2007). Antibodi alami (innate IgM) dikenal sebagai suatu antibodi yang dapat dibentuk sebelum terjadi infeksi ataupun paska vaksinasi. Tingginya avidity IgM menyebabkan IgM dapat mendeteksi dan mengikat antigen kurang reaktif yang sering dijumpai (Boes, 2000). IgM alami merupakan kelas IgM yang telah di deteksi pada manusia dan tikus (Countinho et al., 1995; Haury et al., 1997; Mouthon et al., 1995). IgM alami sebagian besar di hasilkan tanpa adanya eksposur antigen eksogen, hal ini di buktikan dengan ditemukan pada tikus bebas antigen dan
manusia yang baru lahir (Avrameas S, 1991; Casali et al., 1996; Pereira et al., 1986). IgM alami telah diketahui hanya dapat mengenali self-antigen, sehingga terdeteksi dengan kadar rendah untuk infeksi mikroba (Notkin, 2004), akan tetapi saat ini sejumlah penelitian menujukkan bahwa natural IgM dapat mengikat sejumlah mikroba patogen (Ochsenbein et al., 2000; Briels et al., 1981; Gobert et al., 1988; Boes et al., 1998; Baumgarth et al., 2000). Selain IgM sebagai antibodi alami, IgM dapat muncul sebagai akibat telah terjadi paparan imunogen. Berdasarkan hal itu IgM dikelompokkan menjadi dua kelas IgM yaitu innate sel B1 dan adaptive sel B2. Kadar IgM dalam serum dapat muncul karena tubuh terekspose secara akut oleh
sebuah imunogen atau patogen sebagai respon imun primer (Schroeder dan Cavacini, 2010).
IgM telah diketahui sebagai sebuah kekebalan terhadap infeksi sistemik, terutama terhadap virus (Diker, 2005; Mendonsa, 2011). Sapi bali yang dipelihara di Nusa Penida telah diketahui bebas penyakit Jembrana dan SE (Septicemia Epizootica) (Suwiti et al., 2009) yang mana sapi bali sangat rentan terhadap penyakit tersebut. Sehingga kemungkinan adanya kadar IgM pada serum sapi bali di Nusa Penida bukan disebabkan oleh infestasi penyakit akibat virus, melainkan dapat diasumsikan bahwa terdeteksinya kadar IgM pada serum sampel sapi bali disebabkan oleh parasit seperti infestasi ektoparasit ataupun endoparasit. Hal itu didukung karena Indonesia termasuk kedalam wilayah tropis yang memiliki kelembaban nisbi tinggi dan merupakan tempat yang cocok untuk tumbuh berkembangnya ektoparasit ataupun endoparasit. Indraswari et al, (2017) melaporkan bahwa sapi bali di Nusa Penida terinfeksi oleh protozoa gastrointestinal. Studi sebelumnya Batan et al., (2001) melaporkan bahwa sapi bali dapat terinfeksi oleh ektoparasit salah satunya adalah Demodex bovis.
Infeksi parasit dapat mempengaruhi sirkulasi dari IgM dan IgG meskipun telah terjadi respon oleh IgE oleh tubuh (Arlian, 1996), sehingga ketika terjadi infestasi parasit pada sapi bali dapat menyebabkan terstimulasi dan meningkatnya kadar IgE, IgM dan IgG (Morsy et al., 1993). Studi lain melaporkan bahwa infestasi arthropoda pada sapi dapat merubah nilai hematologi yang secara spesifik terjadi peningkatan pada eusinofil dan peningkatan nilai limfosit. Hal ini yang mempengaruhi dalam produksi IgM (Raut et al., 2008; Wikel, 1985).
Kadar IgM serum sapi bali yang berada di Nusa penida pada penelitian ini sangat bervariasi dan memiliki rentang nilai yang jauh berkisar dari 2.565 ng/mL -8.834
ng/mL. Hal itu diasumsikan karena terjadi perbedaan kondisi lingkungan dan pemeliharaan. Studi sebelumnya Bayram et al., (2016) mendeteksi kadar IgM pada sapi Holstein Friesian juga memiliki kadar yang rendah serta bervariasi karena di pengaruhi oleh kondisi lingkungan dan pemeliharaan. Selain itu kemungkinan bisa juga diakibatkan oleh perbedaan jumlah antigen yang masuk di setiap individu sapi bali yang dapat mempengaruhi kadar IgM. Telah diketahui IgM memiliki waktu paruh yang relatif singkat dalam serum, kira-kira 28 jam, pada tikus normal dengan tidak adanya antigen (Viera dan Rajewsky, 1998). Diasumsikan bahwa produksi IgM berkurang sejak respon maturasi dari sel B. Akan tetapi, kejadian ini tidak selalu menjadi sebuah kasus dan menunjukkan bahwa respon IgM dapat dipertahankan untuk waktu yang lama setelah infeksi atau imunisasi dikarenakan sel B yang berumur panjang (Racine dan Winslow, 2009). Pada studi lain juga melaporkan bahwa IgM alami ada dalam serum dengan titer yang rendah dan berkonstribusi dalam kekebalan awal sebelum timbulnya respon humoral adaptif (Zhou et al., 2007; Haas et al., 2005; Ehrenstein et al., 1998; Fearon et al., 1996).
Secara geografis wilayah, kadar IgM serum sapi bali dipelihara di dataran rendah (4.837±1.385 ng/mL) lebih tinggi daripada di dataran tinggi (4.761±1.353 ng/mL) meskipun juga tidak berbeda nyata secara statistik (P>0.05). Pada penelitian ini bertolak belakang dengan beberapa penelitian yang menyebutkan bahwa kadar IgM sapi di dataran tinggi lebih tinggi daripada di dataran rendah karena berkaitan dengan kondisi lingkungan yang berbeda pada dataran tinggi dan dataran rendah (Mazzullo et al., 2014). Bertolak belakang juga dengan studi sebelumnya yang melaporkan bahwa pada dataran rendah cenderung memiliki tingkat suhu udara yang tinggi, sehingga memudahkan untuk terjadinya stress pada sapi akibat kombinasi kondisi lingkungan yang menyebabkan
suhu lingkungan menjadi lebih tinggi daripada zona suhu nyaman pada sapi (thermoneutral) (Amstrong, 1994). Terpaparnya sapi dengan kondisi suhu lingkungan yang tinggi menstimulasi mekanisme termoregulasi dan penurunan tingkat metabolisme, nafsu makan dan produktivitas (Abdelatif dan Alameen, 2012).
Pada penelitian ini dapat diasumsikan bahwa tingginya kadar IgM serum sapi bali di dataran rendah dibandingkan di dataran tinggi karena faktor stress akibat kondisi lingkungan ataupun sapi bali telah terinfeksi oleh beberapa penyakit salah satunya adalah parasit. Telah diketahui bahwa infeksi parasit dapat mempengaruhi tingginya kadar IgM serum (Morsy et al., 1993). Telah diketahui bahwa sapi bali juga rentan terhadapa parasit serta di Nusa Penida telah terjadi beberapa kasus penyakit akibat infeksi parasit (Batan et al., 2001; Indraswari et al., 2017). Diketahui juga bahwa Nusa Penida termasuk lahan kritis dan hampir sepanjang tahun dilanda kekeringan sehingga memberikan dampak pada tingkat stress populasi ternaknya. Secara geografis Nusa Penida merupakan kawasan lahan kering, berbukit, dan berzona iklim F yakni dengan distribusi 4 bulan hujan dan 8 bulan kemarau (BPS Klungkung, 2016). Stress dengan kondisi lingkungan dapat merubah dan menurunkan fungsi imun pada sapi dengan perubahan imunitas seluler dan imunitas humoral yang memiliki dampak signifikan pada imunokompetensi (Carroll dan Forsberg, 2007). Penurunan fungsi imun akibat pengaruh kondisi lingkungan dapat menyebabkan penurunan respon imun baik respon secara selular ataupun humoral pada sapi.
Kriteria jenis kelamin pada penelitian ini juga mempengaruhi terhadap kadar IgM serum sapi bali. Kadar IgM serum sapi bali betina (5.018±1.370 ng/mL) lebih tinggi daripada sapi bali jantan (4.477±1.290 ng/mL) meskipun secara statistik tidak signifikan atau tidak berbeda nyata (P>0.05). Studi sebelumnya telah
melaporkan bahwa jenis kelamin memiliki pengaruh signifikan pada kadar IgM serum pada sapi dalam masa penyapihan, dimana sapi betina memiliki kadar konsentrasi tinggi dibandingkan sapi jantan (Akbulut et al., 2003).
Sapi bali betina mengandung hormon estrogen lebih banyak dibandingkan sapi jantan. Hormon estrogen ini merupakan aktifator dari respon imun. Adanya hormon estrogen menyebabkan sel-sel respon imun teraktivasi sehingga lebih tanggap terhadap antigen yang masuk ke dalam tubuh (Rasyid et al., 2008). Selain itu, didukung pula penelitian oleh Mirzadeh et al, (2010) dengan membandingkan salah satu kriteria penelitian terhadap jenis kelamin yaitu jantan dan betina terhadap gambaran hematologi yang menujukkan bahwa sapi betina memiliki nilai leukosit yang tinggi dibandingkan dengan sapi jantan. Hal itu dapat diasumsikan bahwa respon sapi bali betina lebih tanggap dibanding sapi bali jantan ketika terjadi adanya infeksi, sehingga kemampuan produksi IgM lebih cepat dan memiliki kadar yang lebih tinggi jika dibandingkan dengan jantan.
Pada kriteria umur, kadar IgM serum sapi bali dewasa (4.869±1.417 ng/mL) lebih tinggi daripada kadar IgM serum sapi bali muda (4.707±1.305 ng/mL) meskipun tidak terdapat perbedaan nyata secara statistik (P>0.05). Hal itu sejalan dengan studi sebelumnya oleh jazek et al., (2012) bahwa sapi di usia muda memiliki kadar immunoglobulin lebih rendah dibandingkan dengan usia dewasa. Hal itu dipengaruhi oleh transfer pasif kolostrum pada saat usia muda, sedangkan saat usia dewasa sapi sudah terjadi auto-sintesis immunoglobulin (Erhard et al., 1999). Akan tetapi faktor lain seperti adanya infeksi oleh parasit juga dapat mempengaruhi kadar IgM dan IgG serum sapi (Arlian, 1996).
SIMPUALAN
Simpulan
Sebanyak 54 sampel terdeteksi adanya kadar imunoglobulin M (IgM) dengan nilai
yang bervariasi berkisar dari 2.565 ng/mL -8.834 ng/mL. Kadar IgM serum sapi bali di dataran rendah (4.837±1.385 ng/mL) lebih tinggi daripada di dataran tinggi (4.761±1.353 ng/mL), kadar IgM serum sapi bali dewasa (5.018±1.370 ng/mL) lebih tinggi daripada muda (4.477±1.290 ng/mL) dan kadar IgM serum sapi bali betina (4.869±1.417 ng/mL) lebih tinggi daripada jantan (4.707±1.305 ng/mL). Pada setiap kategori seperti geografis wilayah (dataran tinggi dan dataran rendah), jenis kelamin (betina dan jantan), umur (muda dan dewasa) memiliki nilai rerata P>0,05.
Saran
Perlu dilakukan penelitian lanjutan menggunakan jumlah sampel yang lebih banyak dengan kriteria bervariasi dan membandingkan sampel serum dengan sapi bali yang dipelihara di luar daerah Nusa Penida supaya dapat mengetahui perbandingan bagaimana kadar IgM serum sapi bali di Nusa Penida dan di luar daerah Nusa penida.
UCAPAN TERIMA KASIH
Penulis mengucapkan terimakasih kepada Kepala Balai Besar Veteriner, Denpasar yang telah memberikan izin dan fasilitas untuk melakukan penelitian ini.
DAFTAR PUSTAKA
Abbas KA, Lichtmant AH, Pillai S. 2007. Cellular and Molecular Immunology. 6th Ed. WB Saunders Company.
Abdelatif AM, Alameen AO. 2012. Influence of season and pregnancy on thermal and haemotological responses of crossbred dairy cows in a tropical environment. Global. Vet., 9: 334-340.
Akbulut O, Bayram B, Yanar M. 2003. Serum immunoglobulin concentration of Brown Swiss and Holstein Friesian calves and their relationship with growth characteristics. Ataturk. Univ. Ziraat. Fak. Derg., 34(2): 157-159.
Armstrong DV. 1994. Heat stress interaction with shade and cooling. J. Dairy. Sci., 7: 2044-2050.
Arlian LG. 1996. Immunology of scabies-The Immunology of Host-Ectopara-sitic Arthropod Relationships. CAB International.Wallingford, UK.
Antara PATK, Suwiti NK, Apsari IAP. 2017. Prevalensi nematoda gastro intestinal bibit sapi bali di Nusa Penida. Bul. Vet. Udayana, 9(2): 195-201.
Avrameas S. 1991. Natural autoantibodies: from ‘horror autotoxicus’ to ‘gnothi seauton’. Immunol. Today., 12(15): 4-9.
Baumgarth N, Herman OC, Jager GC, Brown LE, Herzenberg LA, Chen J. 2000. B-1 and B-2 cell-derived
immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J. Exp. Med. 192(2): 71-80.
Batan IW, Wiyanti NWS, Wirat P. 2001. Pola penyebaran lesi demodekosis sapi bali dan efektifitas pengobatan doramectin. J. Vet., 2(2): 49-54.
Baumgarth N, Herman OC, Jager GC, Brown LE, Herzenberg LA, Chen J. 2000. B-1 and B-2 cell-derived
immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J. Exp. Med., 192(2): 71-80.
Bayram B, Aksakal V, Turan I, Demir S, Mazlum H, Cosar I. 2016. Comparison of immunoglobulin (IgG, IgM) concentration in calves raised under organic and conventional condition. Indian J. Anim. Res., 50(6): 995-999.
Boes M, Prodeus AP, Schmidt T, Carroll MC, Chen J. 1998. A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection. J. Exp. Med., 188(238): 1-6.
Boes M. 2000. Role of natural and immune IgM antibodies in immune responses. Mol. Immunol., 37: 1141-1149.
Briles DE, Nahm M, Schroer K, Davie J, Baker P, Kearney J. 1981. Antiphosphocholine antibodies found in normal mouse serum are protective against intravenous infection with type
3 Streptococcus pneumoniae. J. Exp. Med., 153: 694-705.
Carroll JA dan Forsberg NE. 2007. Influence of stress and nutrition on cattle immunity. Vet. Clin. North Am. Food Anim. Pract., (23): 105-149.
Casali P, Schettino EW. 1996. Structure and function of natural antibodies. Curr. Top Microbiol. Immunol., 210: 167-179.
Coutinho A, Kazatchkine MD, Avrameas S. 1995. Natural autoantibodies. Curr. Opin. Immunol., 7: 812-818.
Damayanti R. 2016. Penyakit Malignant Catarrhal Fever di Indonesia dan upaya pengendaliannya. Wartazoa, 26(3): 103-114.
Departemen Pertanian. 2006. Peraturan Menteri Pertanian tentang Sistem Perbibitan Ternak Nasional, Direktur Jenderal Peternakan. Jakarta.
Diker KS. 2005. Immunoloji. Medisan Yayin Serisi: 37, Ankara.
Ehrenstein MR, O’Keefe TL, Davies SL, Neuberger MS. 1998. Targeted gene disruption reveals a role for natural secretory IgMin the maturation of the primary immune response. Proc. Natl. Acad. Sci. USA., 95(100): 89-93.
Erhard MH, Amon P, Younana M, Ali Z, Stangassinger M. 1999. Absorption and synthesis of immunoglobulins g in newborn calves. Reprod. Domest. Anim., 34: 173-175.
Fearon DT, Locksley RM. 1996. The instructive role of innate immunity in the acquired immune response. Science, 272(5): 1-4.
Gobet R, Cerny A, Ruedi E, Hengartner H, Zinkernagel RM. 1988. The role of antibodies in natural and acquired resistance of mice to vesicular
stomatitis virus. Exp. Cell. Biol., 56(1): 75-80.
Haas KM, Poe JC, Steeber DA, Tedder TF. 2005. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity, 23: 7-18.
Handriwirawan E dan Subandriyo. 2004. Potensi keragaman sumberdaya genetik sapi bali. Wartazoa, 14(3): 50-60.
Haury M, Sundblad A, Grandien A, Barreau C, Coutinho A, Nobrega A. 1997. The repertoire of serum IgM in normal mice is largely independent of external antigenic contact. Eur. J. Immunol., 27(15): 57-63.
Indriawati, Margawati ET, Ridwan M. 2013. Identifikasi Virus penyakit jemberana pada sapi bali menggunakan penanda molekular Gen env SU. Berita Biologi, 12(2): 211-216.
Indraswari AASI, Suwiti NK, Apsari IAP. 2017. Protozoa gastrointestinal: Eimeria auburnensis dan Eimeria bovis menginfeksi sapi bali betina di Nusa Penida. Bul. Vet. Udayana, 9(1): 112116.
Iskandar. 2011. Performa reproduksi sapi po pada dataran rendah dan dataran tinggi di Provinsi Jambi. J. Ilmiah Ilmu-ilmu Peternakan, 14(1): 51-61.
Jazek J, Malovrh T, Klinkon M. 2012. Serum Immunoglobulin (IgG, IgM, IgA) concentration in cows and their calves. Acta Agric. Slovenica, 3: 295298.
Korhonen H, Kaartinen L. 1995. Changes in the composition of milk induced by mastitis. The Bovine Udder and Mastitis. Iyvaskyla, Finland:
Gummerus Kirjapaino Oy. Pp. 76-82.
Kresno SB. 2001. Imunologi: Diagnosis dan Prosedur laboratorium Edisi 4. Jakarta: Universitas Indonesia Press.
Mazzullo G, Rafici C, Commarata F, Caccamo G, Rizzo M, dan Piccione G. 2014. Effect of different environmental conditions on some haematological parameters in cow. Ann. Anim. Sci. 14(4): 947-954.
Mendonsa KM. 2011. Factors affecting passive transfer in neonatal calves. Dairy Science Department. California Polytechnic State University.
Mirzadeh KH, Tabatabaei S, Bojarpour M, Mamoei M. 2010. Comparative study of hematological parameters according
strain, age, sex, physiological status and season in iranian cattle. J. Anim. Vet. Adv., 9(16): 2123-2127.
Morsy TA, Kenawi MZ, Zohdy HA, Abdalla KF, Fakahany AFE. 1993. Serum immunoglobulin and
complement values in scabietic patients. J. Egypt. Soc. Parasitol. 23: 221-228.
Mouthon L, Nobrega A, Nicolas N, Kaveri SV, Barreau C, Coutinho A. 1995. Invariance and restriction toward a limited set of self-antigens characterize neonatal IgM antibody repertoires. Proc. Natl. Acad. Sci. USA., 92(38): 3943.
Notkins AL. 2004. Polyreactivity of antibody molecules. Trends Immunol., 25: 174-179.
Ochsenbein AF, Zinkernagel RM. 2000. Natural antibodies and complement link innate and acquired immunity. Immunol. Today, 21: 624-630.
Pane I. 1991. Produktivitas dan breeding sapi Bali. Pros. Seminar Nasional Sapi Bali. Ujung Pandang. 2-3 September 1991.
Pereira P, Forni L, Larsson EL, Cooper M, Heusser C, Coutinho A. 1986. Autonomous activation of B and T cells in antigen-free mice. Eur. J. Immunol., 16(68): 5-8.
Putro PP. 2004. Pencegahan, Pengendalian dan Pemberantasan Penyakit Hewan Menular Strategis Dalam
Pengembangan Usaha Sapi Potong. Lokakarya Nasional Sapi Potong, Pp. 22-26.
Racine R dan Winslow GM. 2009. IgM in microbial infection: Taken for granted. Immunol. Lett., 125: 79-85.
Rasyid R, Yanwirasti, Nasrul E. 2008. Pengaruh Esterogen Terhadap Aktifitas Sel Makrofag dalam Menfagosit Candida albicans Secara Invitro. Majalah Ked Andalas, 1(32): 79-87.
Raut PA, Sonkhusale VG, Khan LA, Nakade MK, Pagrut NS, Boodkhe AM. 2008. Haematological changes in cattle
associated with arthropods infestation. Vet. World, 1(11): 338-339.
Sampurna IP. 2012. Analisis regresi nonlinear terapan dengan spss. Pelawa Sari. Denpasar.
Sampurna IP, Nindhia TS. 2008. Analisis Data dengan SPSS dalam Ran cangan Percobaan. Udayana University Press.
Sayaka B. 2012. Pengembangan Perbenihan Sapi Potong dan Perananya Dalam Pencapaian Swasembada Daging Sapi. Forum Peneliti Agro Ekonomi, 30(1): 59-71.
Schroeder HW dan Cavacini L. 2010.
Structure and function of
immunoglobulins. J. Allergy Clin. Immunol., 125(202): 1-24.
Surhayati S, Hartono M. 2015. Pengaruh manajemen peternakan terhadap efisiensi reproduksi sapi bali di Kabupaten Pringinsewu Provinsi Lampung. J. Penelitian Pertanian Terapan, 16(1): 61-67.
Suwiti NK. 2009. Fenomena jembrana disease dan bovine immunodeficiency virus pada sapi bali. Bul. Vet. Udayana. 1(1): 21-25.
Telupere FMS dan Katipana NGF. 2014. Pengaruh ketinggian tempat dan sistem pemeliharaan terhadap korelasi genetik. J. Nukleus Peternakan, 1(1): 1-6.
Vieira P, Rajewsky K. 1988. The half-lives of serum immunoglobulins in adult mice. Eur. J. Immunol., 18: 313-316.
Wikel SK, 1985. Effects of tick infestation on the plaqueforming cell response to a thymic dependant antigen. Ann. Trop. Med. Parasitol., 79: 195-198.
Zagorska J, Ciproviča I, Miķelsone V. 2007. Baktericīdo vielu un antivielu satura izvērtējums dažādās
lauksaimniecības sistēmās turēto govju pienā. Latvijas Lauksaimniecības Universitātes Raksti, 18(313): 45-50.
Zhou ZH, Tzioufas AG, Notkins AL. 2007. Properties and function of polyreactive antibodies and polyreactive antigenbinding B cells. J. Autoimmun, 29(21): 9-28.
93
Discussion and feedback