Response of North Pacific Seastar Asterias amurensis To Food Attractant (Betaine)
on
JURNAL BIOLOGI XVI (2) : 52 - 56
ISSN : 1410 5292
RESPONSE OF NORTH PACIFIC SEASTAR Asterias amurensis TO FOOD ATTRACTANT (BETAINE)*)
Deny Suhernawan Yusup
Department of Biology Faculty of Math and Natural Science University of Udayana
Email: dsyusup@yahoo.com
INSTISARI
Penelitian tentang respon hewan bintang laut Pasifik Utara telah dilakukan di Tasmania Aquaculture and Fisheries Institute (TAFI) Taroona Hobart Tasmania. Tujuan penelitian ini untuk mengetahui respon Asterias amurensis terhadap stimulan (asam amino Betaine) dan ambang konsentrasi yang direspon serta orientasi pemangsaan (NGDR dan kecepatan merayap). Tujuh konsentrasi (Molar) larutan Betaine yang ditandai pewarna rhodamin digunakan dalam penelitian ini yaitu 0 M (kontrol), 10-1 M; 10-3 M; 10-4 M; 10-5 M; 10-6 M; 10-7 M; 10-8 M. Kecepatan arus yang digunakan adalah 1,1 cm/dt. Respon direkam dengan kamera video dan dianalisa dengan program MOCHA. Hasil analisa statistik menunjukkan bahwa respon maksimum ditunjukkan pada konsentrasi 10-5 M (P: 0,05, IDF= 3,843). Kosentrasi ambang diduga antara 10-7 M dan 10-8 M. Hasil penelitian juga menunjukkan bahwa tidak ada perbedaan respon bintang laut kecil dengan yang besar. Hasil analisa statistik juga menunjukkan bahwa NGDR dan kecepatan merayap tidak ada perbedaan diantara konsentrasi Betaine.
Kata kunci: Chemosensory, Asterias amurensis, Perilaku Makan, Foraging, Sensitivitas
ABSTRACT
A study on the response of North Pacific Sea star was undertaken at Tasmania Aquaculture and Fisheries Institute (TAFI) Taroona Hobart Tasmania. The objectives of the observation were to observe response of Asterias amurensis to various of feeding attractant (Betaine-amino acid) concentration, the threshold concentration to which A. amurensis respond; feeding orientation (NGDR) and crawling spped. Seven molars concentration of feeding attractant dyed with rhodamine were applied i.e. 0 M (control), 10-1 M; 10-3 M; 10-4 M; 10-5 M; 10-6 M; 10-7 M; 10-8 M. The experiment was carried out in a simulation tank with undirectional flow i.e. 1,1 cm/s. Feeding response was recorded with video camera and quantitatively analyzed by means of image analysis software i.e. MOCHA. The statistical analysis results indicate that the maximum response was shown at 10-5 M (P: 0,05), IDF= 3,843) and the threshold concentration was thought at between 10-7 M and 10-8. It was also shown that there was not significant difference on the response of small and large size individuals, except at 10-1M concentration. The were no significant difference on both NGDR and crawling speed.
Key words: Chemosensory, Asteias amurensis, Feeding, Foraging, Sensitivity
INTRODUCTION
The marine environment is crowded with a variety of molecules at various concentrations, resulting from processes occurred in the water and in or on the seabed. These molecules could act as stimulant for the activities of many animals. In the aquatic environment, the optimization of foraging by those creatures relying on an olfactory sensory organ, which could be correlated with the ability to sense distant food qualitatively and quantitatively from the cues received (Zimmer-Faust, 1987). Foragers living in an aquatic habitat should be able to distinguish the alteration occurred in a particular substance from the background chemical mixture diluted in the water. Such ability is related to energy gained, where the energy obtained is determined by the sensitivity of a forager to such changes in orienting and locating the resources.
In understanding the relationship between chemo-
reception and ecology, it is necessary to determine the minimum concentration at which an animal can detect a particular molecule in water. Studies on threshold concentration have been conducted by many researchers for example: Pearson et al. (1979) and Mackie and Shelton (1972). Nevertheless, there are some points which should be noted such as, whole food extract is more attractive than a single or combination of amino acids as demonstrated by Pearson et al. (1979) who worked with the Dungeness crab (Cancer magister). Moreover Mackie and Shelton (1972) who studied lobster (Homa-rus gammarus) also found that no single component was as attractive to lobster as the whole prey. In asteroids, McClintock et al. (1984) revealed that Luidia clathrata responded strongly to components often found in preferred dietary items. Two of the latest authors and Val-entincin (1985) found that the response varied according to different substances. Reaction of a given stimulant with other substances could influenced the response of a
Manuscript has been presented at International Seminar of MIPA’Net, Denpasar Nopember 2009
given animal (Zafiriou et al., 1972). Zafiriou et al. (1972) demonstrated that the presence of silica gel significantly decreased the response of Asterias vulgaris to oyster homogenate. Other studies demonstrated that a mixture of amino acids was nearly as effective as extract of prey, however when tested alone, the amino acids had only a modest stimulatory capacity. Further investigations of seastar responses to feeding attractants are needed.
The present study used betaine, an amino acid commonly found in fish, crabs, and molluscs as a feeding attractant. It has been demonstrated that betaine is able to elicit a feeding response in the seastar Luidia clathrata (McClintock et al., 1984). The amino acid has also been used in a study of the feeding behaviour of other carnivores such as in Palaemonetes pugio (Carr, 1978) and Macrobrachium rosenbergii (Harpaz and Steiner, 1990).
The aims of this study were to investigate whether Asterias amurensis would be able to locate prey on the surface by tracking an odour plume of betaine solution, and the threshold sensitivity of A. amurensis to this amino acid.
MATERIALS AND METHODS
Betaine solutions dyed with rhodamine were used as the source of attractant. The attractant was consisted of 7 molar concentrations i.e. 10-1 M, 10-3 M, 10-4 M, 10-5 M, 10-6 M, 10-7 M, 10-8 M, and 0 M (rhodamine solution only) as the control. The flow rate applied in this study was similar to the slow velocity used earlier (1.1 cm s-1). The flow rate was the mean obtained by reference to water discharge (Q). This flow rate resulted in a continuous odour plume on the flat surface sediment. Seastars were fed with Paphies and starved for up to 7 days before the trials were commenced.. The seastars were grouped into two size based on the radius of arms i.e. large (R > 33.0 mm) and small (R < 32.5 mm). Crawling-speed and NGDR of seastars were quantified from video analysis. T- tests (2 tailed) were used to test whether crawling speed or NGDR varied between concentrations.
RESULTS
Threshold Concentration
The response of seastars to molar concentration of betaine solution is presented in Fig.1. The responses resulted in a bell-shaped graph with 10-5 M producing the maximum response. The proportion of successful seastars increased from 2.44 % ; in the control to a peak of 58.82% in 10-5M but changed only slightly from 10-4 to 10- 1.
The low response of of seastars to molar concentration of betaine solution in controls indicated that the seastars were not attracted to rhodamine solution, nor to water flow alone. Moreover, the low responses to high molar concentrations suggest that the betaine becomes a repellant at these levels.
The Chi-square analysis indicates that the control (0 M) was significantly different from all concentrations except
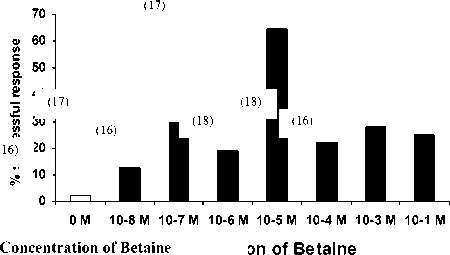
Fig 1. The response of A. amurensis to Betaine solution (flow rate 1.1. cm s-1). The number in the brackets indicates the number of seastar in each trial
10-8 M, while 10-8 was not significantly different from 10-7 M This indicated that the threshold concentration, i.e. the minimum, betaine concentration needed for eliciting feeding behaviour, was between 10-8 M and 10-1 M. The effect of size on response of A. amurensis is presented in Fig. 2. No small seastars were successful in the control and 10-1 M. A chi-square analysis indicated that the only significant difference between small and large was at 10-1M (P= 0.0147)
Feeding Orientation
Crawling speed and NGDR of A. amurensis in response to these betaine concentrations are presented in Figs 3 and 4. Statistical analysis by means of two tailed t-tests (unequal replication) indicated that the crawling speed was not significantly different between concentrations (P > 0.05) except between 10-5 and 10-4 M. There was also non significantly different NGDRs at these concentrations (P > 0.05).
DISCUSSION
Many aquatic organisms rely on waterborne chemical cues and incoming flow to detect their prey using their sensory organs, discriminating food sources and identifying their enemies. As organisms with poorly developed visual sense, seastars has also been demonstrated to rely heavily on waterborne chemical cues. It is thought that seastars are able to interpret the cues for their foraging in particular, though there are conflicting conclusions in the literature about such capability. Conflict may arise because the responses are species specific (Sloan, 1980), or may be because seastars are also responding to such physical factors applied in those studies (Moore and Lepper, 1997).
The present study shows convincingly that A.amurensis is capable of responding to a chemical cue released in the water column, and also is able to locate the source of the cue by tracking the odour plume.
Further, the low numbers of successful’ seastars in the absence of an applied odour (control in the experiments) clearly indicates the significant role of the odour cue and yhis finding supports the conclusion above. It clearly indicates that a successful response is unlikely to be
Small Large
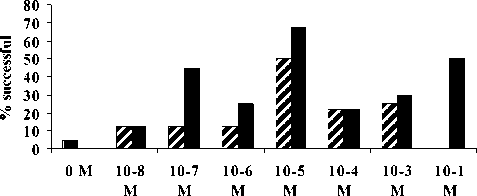
Concentration of Betaine
Fig. 2. The response of small and large Asterias amurensis to different molar concentration of Betaine solution (flow rate 1.1. cm s-1).
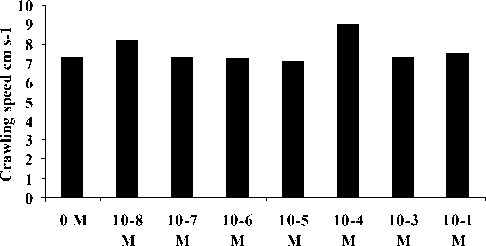
Concentration of betaine
Fig. 3. Crawling speed of A. amurensis to different molar concentration of Betaine solution (flow rate 1.1. cm s-1).
0.8
0.7
0.6
Ctf Q O Z
0.5
0.4
0.3
0.2
0.1
0
IiIIIIII
0 M 10-8 M 10-7 M 10-6 M 10-5 M 10-4 M 10-3 M 10-1 M
Concentration ofBetaine
Fig. 4. NGDR of A. amurensis to different molar concentration of Betaine solution (flow rate 1.1. cm s-1).
due to rheotaxis per se. This finding, to some extent, can be used to counter the arguments of those skeptical about chemosensitivity of seastars, as reported in many studies conducted using flow tanks e.g. Castilla (1972), Zifiriou (1972) and Valentincic (1983).
The findings of the current study agree with the results of a laboratory study conducted by Moore and Lepper (1990) that demonstrating that Asterias forbesi was capable of responding to the presence of an odor, and also be able to locate the odour source in still water. Chemosensitivity responses of A. amurensis in the present study are consistent with the study by Lockhart (1995) on prey preference in flowing water by means
of a Y-maze. If the flow velocity applied in Lockhart’s study i.e. water discharge 3 1 min-1 is converted to mean velocity as used in the present study by calculating the size of the tank, the flow was equal to 0.25 cm s-1 which is fairly similar to the flow applied in the present study (1.1 cm s-1). The results of Lockhart’s study showed the capability of A. amurensis to locate the prey provided in the arms of the maze though they chose the prey inconsistently. Therefore it is unlikely the response was to water current alone. This agrees with the present finding that those seastars tested in the control tank did not show a rheotactic response.
In field conditions, the chemosensitivity of A. amurensis has also been reported by Oliver et al. (1985) who found that A. amurensis was the first predator arriving on damaged substrates resulting from the operation of fishing gear. Though it was not noted, the distance of the seastars from the area, highlights the chemosensitivity of the seastar in perceiving the change of water condition and the capability in tracking the plume under multi-directional flow conditions.
Aquatic predators with poor visual sense rely on information .detected in waterborne plumes. It is known that stimulation of the feeding response will only occur when odor concentration in the water column exceeds the threshold concentration for perception by their sensory organs (Jones 1992, cited in Hara, 1992).
The results of the present study indicated that the response of A. amurensis to chemical cue is concentration related. Response to different betaine concentrations was bell-shaped; the responses increased with increasing concentration only up to some apparent optimal level, and then decreased as the concentration further increased. The low response in low concentration may be because the cue was too weak to elicit a feeding response or it was too low to be detected by the seastar. In contrast, the low response shown to the high concentration may be because high concentrations act as a repellent to the sensory organs of the seastar. A physiological explanation for the bell-shaped response phenomenon is lacking
Similar results have been reported by Carr (1978) for Palaemonetes pugio. It is suggested that response increases as the odour concentration increases. Such results, however, do not entirely agree with the present results and another study on seastars (McClintock and Lawrence, 1986) which describes the response as increasing as the odour concentration increases, but reaching a plateau even though the odour concentration increased. McClintock and Lawrence (1986) found a similar result from a study on fish.
The response-concentration relationship indicates the threshold concentration of betaine to which A. amurensis responded. The threshold is between 10-7 and 10-8 M. Such a high sensitivity enables A. amurensis to detect changes in chemical compounds from the background concentration in the water column that could indicate the presence of their prey. It also allows the seastar to track an odour to its source through the odour plume formed. Such sensitivity helps to compensate for their limitation of movement when they are searching for
prey. Further it implies the potential of A. amurensis as a very dangerous predator that could influence benthic faunal community structure. This is highlighted by their ability to dig up to 10 cm (adult individual) depth in sediments. Therefore A. amurensis could have massive impact on benthic faunal communities in a given habitat.
The threshold concentration to which seastars are attracted seems to be influenced by many factors such as type of substance, and or physiological state i.e. satiation level. The study by Valentincin (1985) on Marthasterias glacialis demonstrated various behavioural thresholds of the seastar for some effective feeding stimuli such as L-proline, cysteine and acetylcholine iodide. Thresholds may be related to differences in chemical configuration of each amino acid and differences in site attachment at the sensory organ. It likely that such differences could result in different responses from the organism. Similarly, such substance-specific threshold responses have also been demonstrated for other aquatic species such as the seastar, Luidia clathrata (McClintock et al., 1984), shrimp, Palaemonetes Pugio (Carr, 1978) and fishes, genus Savelinus (Harra, 1992)
Another factor that should be taken into account in determining threshold concentration is satiation level. It is thought that the threshold concentration decreases as hunger level increases. A study on sablefish (Anoploma fimbria) demonstrated that the reduction of food supplies significantly increases the sensitivity of the fish to dissolved free amino acids in squid extract. However it remains unclear how such a change in physiological state could increase the sensitivity of sensory organs.
The concentration-response showed that 10-5 M betaine resulted in maximum response. This finding agrees with a previous study on the tubefeet response of Asterias amurensis to different molar concentration applied by means of a cotton bud dosed with betaine (pers. obs). It was found that the fastest tube-feet protraction time was shown when the seastars were offered 10-5 M betaine solution. Though betaine has been recognised as a potent amino acid eliciting feeding behaviour in many aquatic organisms including seastars, in the current study betaine could only motivate modest responses of A. amurensis. Such modest stimulatory capacity of a solution of betaine alone was also shown in the study by Carr (1978) on Palaemonetes pugio. But Carr (1978) also demonstrated that a mixture of amino acids was nearly.as effective as an extract of intact prey such as sea urchin, oyster and blue crab. Another study on Lobster (Homarus gammarus) found that amino acids, either as single compounds or in combination, were less attractive than food extract (Lockhart, 1995).
These studies indicated that arousal of a feeding response is a result of the collective response of many sites in the sensory organ to different elicitor substances contained in a given food source, and is not the result of a response to a single compound per se. Thus laboratory responses to simple solutions are unlikely to be equivalent to the natural response under field conditions. Though there is a difference in efficiency between single compounds and mixtures, the use of
mixtures of amino acids or prey extract could still mimic the synergistic and suppressive interaction that could occur among individual substances, as suggested by Zaimmer-Faut et al. (1984) and Carr and Derby (1986). The study by Zafiriou et al. (1972) demonstrated such suppressive interaction. This study showed that the addition of particulates significantly decreased the effectiveness of the stimulus. In contrast, a study by Carr et al. (1977) on pigfish (Orthopristis chrysopterus) and Zaimmer-Faut et al. (1984) on P. pugio demonstrated a synergistic interaction that increased the effectiveness of the amino acids involved.
It has been widely recognised that waterborne chemical signals play a direct role in guidance of organisms with poor visual ability in searching for an odour source Rochette et al. (1994). The dynamics of odour-plume dispersion in the water column is thought to affect the pattern of tracking including trajectory and speed of an aquatic animal. The current study indicates that there was no clear correlation between pattern of movement (crawling speed and orientation) of A. amurensis and the concentrations of odour tested. It does not support the expectation that there is a positive correlation between concentration and the pattern of movement.
The results of the current study do not agree with those of Carr et al. (1977). Beddingfield and MCClintock (1993) who demonstrated that A. articulatus shows directional pattern in the absence of prey; in contrast when they encounter prey the frequency and magnitude of angular deviation increased. Results also contrasted with those of McClintock and Lawrence (1985) on L. clathrata who found that the seastar changed its response as they encountered an increase in odour concentration by increasing their frequency and magnitude of change in direction of movement. Later the study by Rochette et al. (1994) indicated that Leptasterias polaris showed cross-current behaviour in the absence of odour in the water column. One of the possible reasons for the difference is the structure of the odour plume resulting in uniform responses. In such a continuous plume, the tube-feet of the seastars encountered less temporal variation of signal. In other words the seastars, crawled through a sustained gradient of plume signal. Therefore there was less variation in crawling speed response. Weissburg and Zimmer-Faust (1994) who studied feeding response of blue crab (Callinectes sapidus), found that the crab showed directional responses toward the odour source in less turbulent flow and showed a more indirect trajectory in turbulent flow resulting in smooth and rough odour plumes, respectively. The narrow and straight plume resulting from slow current velocity may cause the tip of the leading arm to move in a limited arc, resulting in a straighter trajectory. The response to some extent conforms to the cross-stream movements of Leptasterias polaris in the absence of odour indicated in the study by Weissburg and Zimmer-Faust (1994). The cross current behaviour is due to a lack of odour signal detected by the tube feet in the leading arm, and it results in much turning behaviour which is thought to be a response serving to increase their chance of locating
odour. McClintock et al. (1984) found that generally the foraging pattern of L. clathrata was directional, however the seastars changed their response, e.g. frequency and magnitude of change in direction of movement, once prey were encountered. The behavior resulted in an individual remaining in a high plume concentration. The study by Dalle (1997) on A. forbesi suggested that the seastars is alteration of stimulus concentration as measured at the tube feet in the tip of each arm to guide them to a food source. In other words the seastars would change their direction of movement to the previous track when faced with a weaker signal.
CONCLUSION
This study indicates that the maximum response of North Pacific Seastar was at 10-5 M and the threshold concentration was thought between 10-7 M and 10-8 M. This study also show that there were not different sensitivity between small and arge individual.
REFFERENCES
Beddingfield, S.D., J.B. McClintock. 1993. Feeding Behavior of the Seastar Astropecten articulatus (Echinodermata: As-teroidea): An Evaluation of Energy-Efficient Foraging in A Soft-Bottom Predator. Mar. Biol. 115 (4): 669-676.
Carr, W.E.S. 1978. Chemoreception in shrimp, Palaemonetes pugio: the Role of Amino Acids and Betaine in Elicitation of A Feeding Response by Extracts. Comp. Biochem. Physiol. 61A: 127-131.
Carr, W.E.S., C. B. Derby. 1986. Behavioral Chemoattractants for the Shrimp, Paleomonetes pugio: Identification of Active Compounds in Food Extract and Evidence of Synergistic Mixture Interaction. Chem, Senses. 11 : 49-64.
Carr, W.E.S., K.M. Blumenthal, J.C. Netherthon. 1977. Chemoreception in the Pigfish, Orthopristis chrysopterus: the Contribution of Amino Acids and Betaine to Stimulation of Feeding Behaviour by Various Extract. Comp. Biochem. Physiol. 58A: 69-73.
Castilla, J.C. 1972. Response of Asterias rubens to Bivalve Prey in a Y-Maze. Mar. Biol. 12: 222-228.
Dalle, J. 1997. Chemosensory Search Behaviour in the Starfish Asterias forbesi. Biol. Bull. 193: 210-212.
Hara, T. J., 1992. Mechanism of Olfaction in Hara, T. J. (ed). Fish Chemoreception. Chapman & Hall. Melbourne. P: 372.
Harpaz, S., J.E. Steiner. 1990. Analysis of Betaine-Induced Feeding Behaviour in the Prawn Macrobrachium rosenbergii (De Man 1879) (Decapoda, Caridae). Crustaceana. 58 (2): 175-185.
Lockhart, S.J. 1995. Feeding Biology of the Introduced Sea Star Asterias amitrensis. Unpublished Honours Thesis. Dep. Zoology Univ. of Tasmania.
Mackie, A.M., R.G.J. Shelton. 1972. A Whole-Animal Bioassay the Determination of the Food Atractants of the Lobster, Homa-rus gammarus. Mar. Biol. 14: 217-221.
McClintock, J.B., J.M. Lawrence. 1984. Ingestive Conditioning in Luidia clathrata (Echinodermata: Asteroidea): Effect of Nutrition Condition on Selectivity, Teloreception and Rates of Ingestion. Mar. Behav. Physiol. 10: 167-181.
McClintock, J.B., J.M. Lawrence. 1985. Characteristic of Foraging in the Softbottom Benthic Starfish Luidia clathrata (Echino-dermata: Asteroidea): Prey Selectivity, Switching Behaviour, Functional Responses and Movement Patterns. Oecologia (Berlin). 66: 291-298,
McClintock, J.B., T.S. Klinger, J.M. Lawrence. 1984. Chemoreception in Luidia clathrata (Echinodermata: Asteroidea): Qualitative and Quantitative Aspects of Chemotactic Responses to Low Molecular Weight Compounds. Mar. Biol.. 84: 47-52.
Moore, P.A., D.M.E. Lepper. 1997. Role of Chemical Signal in the Orientation Behaviour of the Seastar Asterias forbesi. Biol. Bull. 192: 410-417.
Oliver, J.S., R.G. Kvitek, P.N. Slattery. 1985. Walrus Feeding Disturbance: Scavenging Habits and Recolonisation of Bearing Sea Benthos. J. Exp. Mar. Biol. Ecol. 91: 233-246.
Pearson, W.H., P.C. Sugarman, D.L. Woodruff, B.L. 011a. 1979. Threshold for Detection and Feeding Behavior in the Dungeness Crab, Cancer Magister (Dana). J. Exp. Mar. Biol. Ecol., 39:65-78.
Rochette, R., J.F. Hamel, J.H. Himmelman. 1994. Foraging Strategy of the Asteroid Leptasterias polaris: Role of Prey Odour, Current and Feeding Status. Mar. Ecol Prog. Ser. 106: 93-100.
Sloan, N.A. 1980. Aspect of the Feeding Biology of Asteroids. Oceanogr. Mar. Biol. Ann. Rev. 18: 57-124.
Valentincic, T. 1983. Innate and Learned Responses to External Stimuli in Asteroids. In Jangoux, M., J.M. Lawrence (eds) Echinoderm Studies, Vol. 1. Balkema, Rotterdam.
Valentincic, T. 1985. Behavioral Study of Chemoreception in the Seastar Marthasterias glacialis: Structure-Activity Relationship of Lactic Acid, Amino Acid, and Acetylcholine. J. Comp. Physiol. 157: 537-545.
Weissburg, M.J., R.K. Zimmer-Faust. 1994. Odor Plumes and How Blue Crabs Use Them in Finding Prey. J. Exp. Biol. 197: 349-375.
Zafiriou, O., K.J. Whittle, M. Blumer. 1972. Response of Asterias vulgaris to Bivalve and Bivalve Tissue Extracts. Mar. Biol. 13: 137-145.
Zimmer-Faust, R.K. 1987. Crustacean Chemical Perception: Toward A Theory on Optimal Chemoreception. Biol. Bull. 172: 10-29
Zimmer-Faust, RK., J.E. Tyre, W.C. Michel, J.F. Case. 1984. Chemical Mediation of Appetitive Feeding in A Marine Decapod Crustacean: The Importance of Suppression and Synergism. Biol. Bull.
56
Discussion and feedback